





Rediscovering Biology: Molecular to Global Perspectives
Neurobiology Expert Interview Transcript: Richard Huganir, Ph.D.
 Professor of Neuroscience; Investigator, Howard Hughes Medical Institute
Professor of Neuroscience; Investigator, Howard Hughes Medical Institute
Richard Huganir, PhD, is a professor of neuroscience at Johns Hopkins University and an investigator with the Howard Hughes Medical Institute. His laboratory is working to understand the interaction of molecules during long-term potentiation, a neuronal hyperactivity that is thought to underlie learning and memory.
Interview Transcript
Richard Huganir, Ph.D., is a professor of neuroscience at Johns Hopkins University and an investigator with the Howard Hughes Medical Institute. His laboratory is working to understand the interaction of molecules during long-term potentiation, a neuronal hyperactivity that is thought to underlie learning and memory.
Define ‘neurotransmitter.’
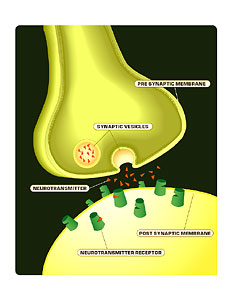
A neurotransmitter is a small molecule chemical that’s released from one neuron and diffuses to a second neuron, and binding to that neuron will transduce a signal between the two neurons.
Define a ‘signal transduction pathway’?
A signal transduction pathway is the pathway by which a neurotransmitter, a signal is transduced. So the neurotransmitter binds to the receptor on the neuron, and this binding is somehow converted into a signal in what we call the postsynaptic neuron.
Define ‘glutamate receptor.’
Glutamate receptors are really the major excitatory receptors in the brain, and they mediate most excitatory neurotransmission. So at an excitatory synapse, glutamate is the neurotransmitters released from the nerve terminal, and it binds to these receptors on the postsynaptic neuron. And these receptors then transduce that signal from the glutamate molecule into an excitatory response. That is, it makes the neuron more excitable, and so triggers the excitability of the postsynaptic neuron.
Are there other receptors for other neurotransmitters?
The glutamate receptors are really exclusively responsive to glutamate. There are many other types of neurotransmitters; the major inhibitory neurotransmitter is a molecule called GABA, Gamma-Amino Butyric Acid.
And this neurotransmitter, just like glutamate, has its own receptors. That is, they are inhibitory neurotransmitters receptors, or GABA receptors, and so GABA binding to these receptors actually decreases the excitability of the neuron, and actually inhibits the neuron from firing or from being excited.
Could you define ‘protein phosphorylation’?
Protein phosphorylation is really one of the most ubiquitous mechanisms for controlling protein function, and it’s a process where enzymes called protein kinases use ATP–adenosine triphosphate–to modify protein structures. So the kinases will take a phosphate molecule from ATP and add it to amino acids, actually serine or tyrosine residues, on a protein.
And so the addition of this chemical group, this phosphate group, to the protein actually makes a negative charge, and that can affect the structure, and also actually regulate the activity or function of that protein.
This is a very ubiquitous system. Protein phosphorylation occurs on most proteins, but it’s also reversible, so you can remove that phosphate. There are enzymes called protein phosphatases that actually cleave the phosphate off and bring the protein back to its sort of more basal state. So, for example, if phosphorylation activates an enzyme or a channel, dephosphorylation will reverse that process and inactivate the enzyme or the channel.
What about its association with memory?
There’s no simple relationship between phosphorylation and memory, but in many systems, phosphorylation of proteins correlates with learning and memory. It correlates with increases in synaptic transmission and connections between neurons, but actually, in other proteins, dephosphorylation may actually increase the connections between neurons. So it really depends on the specific protein that’s getting phosphorylated, and whether phosphorylation is activating or inhibiting that protein, because it’s not a simple one-to-one correlation.
Could you define the ‘CREB protein’?
CREB protein is an activator of gene transcription, and this we know: that learning and memory in many systems requires new gene expression and protein translation. And in these systems, one way to regulate genes is through CREB. So CREB can be activated by protein phosphorylation, and CREB will actually turn on gene production, gene expression, and these genes may be very important for maintaining memory, especially long-term memory.
So are neurons central to brain function?
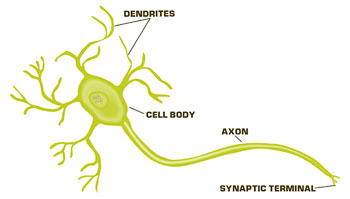
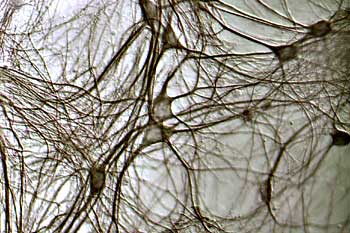
Basically, the brain works by communication between neurons. There are trillions of neurons in the human brain, and it’s the communication between these neurons that make us feel, think, be able to sense, to actually have consciousness. And it’s this continuing communication between neurons that’s important for processing information. But also, the synaptic connections between neurons in our brain are changing all the time, and it’s this change, or what we call “synaptic plasticity,” or changes in synaptic connections that underlie things like learning and memory, or any response to our environments, so the information we take in is processed. We change connections, and then we respond to that environment.
How many synaptic connections are there for each neuron?
Each neuron–we just talked about how there are trillions of neurons in the brain–but each neuron can have up to a thousand different connections, so we’re talking trillions and trillions of synaptic connections, and they are not constant. They are constantly changing.
So synaptic plasticity is extremely important?
Synaptic plasticity is critical for learning, and it’s critical for a plastic, or changing, brain, so to actually process information and come up with something new, you really need to form new connections, and form new associations.
So, for example, when we are learning a new concept, or for a common example, when you are smelling a rose, you smell it, and information, this odor from the rose, goes into your nose and stimulates neurons that go up into your olfactory cortex. At the same time, your eyes see the rose, and that stimulates your retina, which then goes to the visual cortex.
So all this information comes into various regions of the brain at the same time, and timing is very important, so if you’re seeing and smelling the rose at the same time, those two sensory perceptions get correlated and actually form new wiring connections in the brain. What many people say is, “when you fire neurons together, you wire them together,” so firing of the neurons in the olfactory cortex and the visual cortex together wires them together, and makes changes in the synaptic connections, and those synaptic connections form a neuronal network which is the memory of a rose.
Is this a relatively new concept?
It’s really a relatively old concept. Santiago Ramòn y Cajal, one of the pioneers in neuroscience and neuroanatomy back in the late 1800s and early 1900s, actually proposed many of the concepts we have today. He was really a true genius, and he proposed that the idea of a synapse, and changes in synaptic connections, could play an important role in brain function.
Then in the 1950s, D.O. Hebb came up with this idea that by firing together, you wire together; the idea that coincident input into neurons strengthens synapses, and inputs that come in that are not coincident actually weaken synaptic connections, and so these ideas actually have a long history, and go back into the earliest 20th Century.
Is memory a function of neuronal communication?
What most neuroscientists believe is that, to actually encode memories into our brain, you need to set up a network, a neuronal network that can be recalled when you’re remembering something. And so the idea is that there are millions of neurons, and that when you are learning something, you’re setting up a new network through those neurons.
So specific connections–you’re enhancing certain connections between certain neurons, and actually inhibiting connections between other neurons, and in this way you sculpt out a pathway, a neuronal pathway through this network of neurons.
So many of us are interested in what are the sorts of molecular mechanisms that underlie this process of both strengthening and weakening the synaptic connections. So at a molecular level, how do you make neurons communicate better at their points of contact, which are called synapses, and in the same way, how do you reverse this process? How do you weaken a synapse? So this is a bidirectional regulation. You regulate, you increase synaptic connections, between certain sets of neurons, and decrease synaptic connections between other sets of neurons.
Does this form stable connection?
This neuronal network that you set up, that you learn, has to be stable for certainly minutes and days or weeks or even years. Of course, we remember things from our childhood that are 30 to 40, 50, 60 years old. So there have to be very stable mechanisms evolved in setting up these networks, or in increasing the strength of synaptic connections.
One of the mysteries is, how do these connections form and how do they remain stable for that period? And these are huge questions that aren’t going to be answered in the next ten or 20 years, but we’re really getting a good idea of some of the basic mechanisms underlying at least maintaining these type of connections for days, or for a week or so.
Can you cite some examples of these ideas?
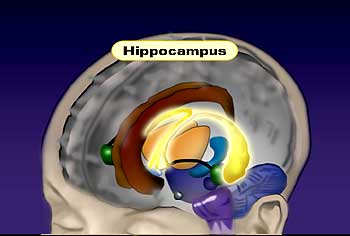
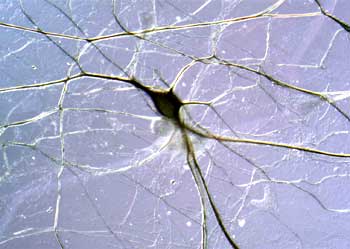
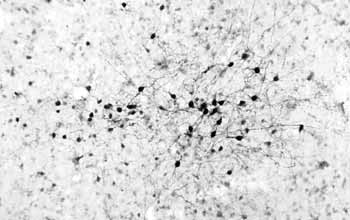
To study these long-term changes in synaptic connections, there are several different model systems that people have used. A lot of people have focused on the area of the brain called the hippocampus, which is a small region of brain, but it’s very crucial for learning and memory.
The importance of the hippocampus in learning and memory was really shown by studying patients who had lesions in that area of the brain, either through stroke or accident, and these patients are very famous, and people have studied them for years. And they essentially have no short-term memories, so these patients, if they have a stroke and this area of the brain is damaged, they’ll remember everything in their childhood and prior to the stroke, but they can’t learn new things. So, for example, they will reintroduce themselves every morning to the doctor that they’ve been seeing for years, and so they can never remember anything new.
People have focused on the hippocampus for this reason, and we try to look at that region of the brain and study its basic properties to find some properties of synaptic transmission that may encode these long-term changes in synaptic transmission.
And there are two phenomena that people study in the hippocampus, one called “long-term potentiation,” and one called “long-term depression,” and these are phenomena where, if you stimulate a neuron and record from another neuron, you can, of course, record synaptic transmission, and if you give very brief, high-frequency stimulus to the one neuron, and record from the second neuron after that high-frequency stimulation, you get a potentiation of the synaptic transmission, and that’s called “long-term potentiation,” because this high-frequency stimulation actually increases the synaptic communication, potentiates it, but this potentiation can last actually days or weeks in the intact animal. So in some way, just a few seconds of stimulation can totally convert the synapse into a more potent synapse. So we’ve been trying to understand at the molecular level what occurs at the synapse.
We know that synaptic transmission occurs through a variety of steps. First, you have an action potential coming down a presynaptic nerve terminal, and this promotes the release of neurotransmitter from the nerve terminal. The neurotransmitter then diffuses across the synaptic cleft and binds to receptors on the postsynaptic neuron. The receptors then transduce the signal of the neurotransmitter, and convey that communication to the second neuron.
To potentiate that process, there are several ways you could do that. One is by, for example, promoting the release of more neurotransmitter. If you release more neurotransmitter, you’re going to get a bigger response in the postsynaptic neuron.
The other way is actually to modify the receptor function, so the receptors are more sensitive to the neurotransmitter, and that way you’ll get a better response to the same amount of released neurotransmitter. And so many people have focused on these two steps. One question is, “Is the release of neurotransmitter regulated during long-term potentiation, or are the receptors or the receptor sensitivity regulated during long-term potentiation?”
And one thing that my lab is really focused on is the receptor systems. We sort of reason that, since the receptors are a convergent point for the neurotransmitters, the neurotransmitters have to bind to the neurotransmitter receptors to convey the signal. So if we could understand how the receptors are regulated, we would understand how synaptic transmission is regulated.
And is the neurotransmitter receptor you’re referring to the glutamate receptor?
We focus on glutamate receptors, because the long-term potentiation is most well studied at excitatory synapses. That’s not to say it doesn’t occur at inhibitory synapses, but certainly, excitatory synapses are the most plastics type of synapses. They are highly modulated, and of course glutamate receptors mediate the response at excitatory synapses, so that’s why we focused on them.
What we’ve focused on primarily is the mechanisms for regulation of receptor function, and, for example, how individual receptors can be regulated. For example, how can you modify receptor to make it more sensitive to the neurotransmitter?
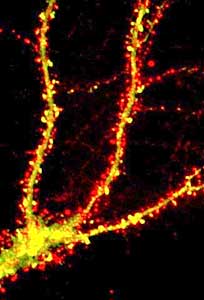
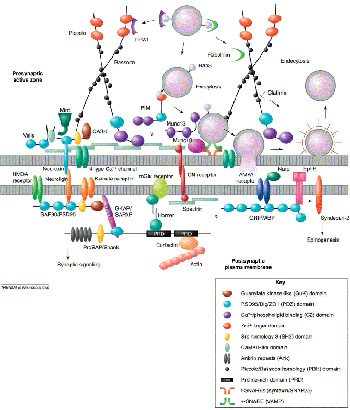
But another way that we’ve been studying receptor function is how the receptor is trafficked or expressed throughout the cell. It’s well known that most receptors are highly concentrated at synapses, so they’re highly concentrated under the nerve terminal that’s releasing glutamate.
But what we now know is that this is a very dynamic process, that you can actually increase or decrease the number of receptors at a synapse very rapidly, within minutes, and of course if you increase the number of receptors of a synapse, you’re going to increase the efficiency of signal transduction, and so you can modify a synaptic communication in that way, too, both by regulating individual receptors, how they individually work and respond to neurotransmitter, but also by regulating the number of receptors at synapse: either up-regulating or down-regulating them to increase or decrease synaptic transmission.
What other chemical changes does the cell use to change receptors?
There are multiple mechanisms for regulating receptor function. One is protein phosphorylation. We discussed protein phosphorylation before. So glutamate receptors themselves get phosphorylated, and phosphorylation of the receptors can affect their function, so for example phosphorylation of the receptors can potentiate their activity, so when the receptor is phosphorylated, it responds better to glutamate, and therefore you get a bigger response at the synapse.
Phosphorylation can also regulate the expression of the reception at the synapse, so phosphorylating the receptor in many cases actually increases the stability of the receptor at the synapse. So you actually have a higher number of receptors at the synapse. So those are two ways of regulating existing synapses, but there’s actually a lot of interest now in the formation of physically new synapses by the formation of new synaptic structures. So excitatory synapses have a unique structure, in that they actually occur on small protrusions from a neuron called “synaptic spines,” and these small protrusions are very dynamic. They can wiggle around, and they can retract or expand out from the dendrites, and so this is another way that a lot of people are looking at these dendritic spines to try to understand what regulates these spine protrusions and the formation of these new spines synapses.
So, for example, during LTP there’s an interest now that during induction of LTP, long-term potentiation, you actually get sprouting of new spines, and these form new synapses, and so not only are you modifying existing synapses by modifying say receptor number or receptor function, you’re also physically making new synapses by what we call sprouting of synaptic spines.
So this involves a structural change in proteins?
During LTP, we found that the glutamate receptors get phosphorylated, and upon this phosphorylation, we found it actually regulates the activity of the receptor, and presumably, this occurs through the addition of phosphate molecules, which somehow affects the structure or confirmation of the receptor and thereby changes its physical properties.
So these receptors are ion channels. When they bind glutamate, they open a pore, or channel, through the membrane and allow ions to flow through the channel. And what we found is that when you phosphorylate the receptors, the receptors actually allow more ions per second go through them, so you’re getting a bigger response. You’re getting more ions passing through the receptor for the same amount of binding. We found the phosphorylation affects the structure and the conformation of the receptor.
And this structural change has an effect on the ion channel’s function?
When the receptor is phosphorylated, it actually passes more ions per second through the ion channel, so for the same amount of neurotransmitter binding to the receptor, you actually get a bigger flux of ions across the membrane.
And what are the effects of dephosphorylation?
Long-term potentiation is one process to increase the connections between neurons, but there’s an opposite process called “long-term depression,” and this is a process whereby you actually inhibit or decrease the connectivity between neurons. And actually this turns out to be in principle, at the molecular level, the reverse of LTP, and so instead of having kinases that phosphorylate the receptors, what we found is that there are protein phosphatases that dephosphorylate the receptor.
So as I mentioned, phosphorylation of the receptor potentiates the receptor function, but during long-term depression that phosphorylated residue on the receptor is dephosphorylated, so you reverse that process and actually inhibit receptor function. So it’s a reversing effect.
And how does this affect memory?
At a simplified level, you could think of long-term depression as forgetting; you’re actually disconnecting neurons. But I think that’s an oversimplification. You need both strengthening and weakening of synapses to sculpt the neuronal circuits, and so both are playing a positive role in setting up neuronal pathways that encode memories. But of course, too, if you want to erase that memory, that neuronal circuit, you would have to disconnect those neurons, and therefore, in that way, you can think of long-term depression and dephosphorylation of the receptors as ‘forgetting.’
So let’s review: what are kinases and phosphatases?
Protein kinases are enzymes that are very ubiquitous in all cell types. But they’re very highly expressed, or present, in neurons. These enzymes take ATP, the energy source in the cell, and use it as a substrate. They cleave off a phosphate molecule from the ATP and covalently add this to the protein, so the enzyme catalyzes or enhances this reaction, and adds the phosphate to the protein. In this case, what we’re talking about are receptors, so the kinase adds the phosphate molecule to the receptor and thereby changes its structure and its function.
Protein phosphatases are again very ubiquitous enzymes. They’re present in most cell types, liver, any cell type, but they’re again very highly expressed in the brain. These enzymes are enzymes that help to catalyze the removal of phosphate from proteins. So once a protein is phosphorylated by kinases, phosphatases are the enzyme that reverse this process and remove the phosphate through a hydrolytic reaction and return the protein to its basal state.
So actually the phosphorylation level of a protein is a balance between kinase activity and phosphatase activity; whichever is more active will win the day, and so if a kinase is much more active than a phosphatase, you’ll have a lot of phosphorylated protein, and vice versa. If the phosphatase is much more active, you’ll have a dephosphorylated, a non-phosphorylated protein.
What role do calcium channels play?
In the process of long-term potentiation, calcium plays a central role. Calcium entry into the postsynaptic cell is what triggers the signal transduction pathways that ultimately result in potentiation of the synapse. And this signal transduction pathway initiated by calcium has been studied for the last 20 years or so, and we know a lot about it. We know how calcium plays this role in inducing long-term potentiation. One of its primary roles is actually activating a protein kinase called “calcium-calmodulin-dependent kinase two,” or CaMKII, as well as others. So calcium triggers the activation of kinases, which then phosphorylate the receptors and potentiate their function. So that’s one of the major steps in regulating and inducing long-term potentiation.
Could you use a gate as a metaphor for the calcium channel?
Glutamate binding to its receptor opens a gate, which allows ions, including calcium, to go across the membrane.
So how can we relate this molecular mechanism of LTP to real-world learning and memory?
The idea that LTP, or long-term potentiation, underlies learning and memory is a very large step. We’re actually studying a basic cellular mechanism in the brain. It’s a very fascinating mechanism, but still, directly relating it to learning and memory has been very difficult. And the best researchers can do is really to correlate learning and memory with long-term potentiation.
The way many of us have done this now is to try to dissect long-term potentiation at a molecular level, and then manipulate that molecular system and ask the question: “Does that manipulate learning and memory?” And of course the way that we have done this now is through genetic methods in mice, where you can study the behavior of the mice, study their learning behaviors, and using the power of molecular biology, we can actually genetically modify mice to make what we call ‘designer mice,’ and for example mutate certain genes that we think are in the pathway.
One of the classic studies to look at this was to knock out this CaMKII gene that I talked about. We know that this kinase is critical for LTP, and so the idea was to actually genetically delete it in mice, to make a mouse that has no CaMKII, and then, first of all, ask the question: Does it still have LTP? And it turned out that it did not have LTP.
But then secondly, it was important to study its learning and memory capabilities. And it turned out that these mice have very poor learning and memory, so at least at that level, the correlation held that if you eliminated a gene that we know is important for long-term potentiation, it also dramatically affected the mouse behavior, its learning and memory.
We are now doing this more and more subtly. We’re making more subtle mutations. For example, one study that we’ve recently done is to actually mutate the phosphorylation sites in the gene for these glutamate receptors, and what we’ve been able to do is mutate the glutamate receptor gene so it can no longer be phosphorylated at the place that is normally phosphorylated during LTP.
So what we’ve made is a mouse that is totally normal, has a normal receptor gene, but this receptor gene can no longer be phosphorylated on those sites. And now we’re studying their behavior, and it turns out that these mice actually learn fairly well, but they forget very quickly. Once you train them in a task, they will remember for two to four hours, but after about eight hours they start forgetting the task, compared to wild-type animals, which will remember the task for several days.
So again, we think that phosphorylation of receptors is very important for LTP, and we’ve found that mutating those phosphorylation sites inhibited the formation of LTP in these mutant mice but also had effects on behavior, on learning and memory. In trying to understand the regulation of receptor function, one obvious way was to study how the receptors are phosphorylated, and how that regulates the properties. But we knew, as I mentioned earlier, that the numbers of receptors present can be modified at the synapse, so we really wanted to understand how that occurs. It’s really pretty remarkable that the receptors are highly concentrated right at a very specific area on the neuron, so the question is: How does the receptor know where to go, and once it gets there, what stabilizes it at that point of contact?
And so what we’ve done is to try to identify other gene products, other proteins that actually bind or physically interact with receptors. These other molecules may play a role, for example, in guiding structural proteins out from the cell body, where they’re made, to the synapse, where they’re active. And then once they get to the synapse, they bind to the receptor and actually help stabilize the synapse.
So we did a comprehensive screen to try to identify proteins that bind to the receptors. We have isolated several proteins that specifically bind to the receptors, and one of these is called GRIP, or “glutamate receptor interacting protein,” and this protein we now think is involved in promoting the export of the receptor into the dendrite and into the synapse, and so GRIP would play a positive role. It actually would promote the insertion of receptors into the synapse and would promote, or potentiate, synaptic transmission.
In contrast to GRIP, we’ve also found other proteins, for example, a previously identified protein called PICK1, and we think this protein actually has the opposite effect. It actually promotes the removal of receptors from the synapses. So we think what happens is that the PICK1 protein, when it binds to a receptor, actually helps actively remove the receptor from the synapse, so PICK1 would play a negative role, in decreasing the number of receptors at the synapse and therefore decreasing the efficiency of synaptic transmission.
Why would receptors be removed?
One possibility during long-term potentiation is that you add receptors to the synapse to increase synaptic transmission. We also think that the opposite occurs during LTD, long-term depression; that during LTD, you actually remove receptors from the synapse and therefore decrease the synaptic transmission.
So we think that PICK plays an active role in LTD, that during long-term depression, PICK will bind to the receptor, bring it into the interior of the cell and therefore decrease the number of receptors at the synapse.
Would you say that the 1990s were the decade of the brain?
I think the 1990s were a decade of incredible progress in neuroscience, but I think it’s only the tip of the iceberg. We’ve come a long way, especially in the molecular analysis of synaptic function. But really I think we’re at the beginning of a whole new field of really how the brain works. I mean if you think about the brain, it’s such a complex organ, and although we understand how synapses work to a certain degree, we have very little idea how higher brain processes occur, and I think probably that this is going to be the century of the brain, and we will really begin to understand how higher brain functions occur at what we call a “systems-level” in neuroscience, and possibly even things such as consciousness. To try to understand consciousness is really a final goal of neuroscience, and we’ll see what happens in the next hundred years.
What new techniques do you think will have a major impact?
The brain actually works through electrical signals, so to really measure neuronal function you have to electrically record the activity of these neurons. And the way we do that is by a relatively new technique called “patch clamping,” where you actually take a glass electrode and place it up against the cell and actually sort of stick it on to the cell, so you can measure electrical currents going from the inside to the outside of the cell.
And in this way, we can record synaptic transmission between neurons. So, for example, when synaptic transmission occurs, you can actually measure small deflections in the membrane potential across the neuron’s membrane. And we use this technique to really measure synaptic transmission. And this is of course how you measure the strength of connection, by the change in the potential across the membrane of the neuron.
What is the substance in the pipette?
When you use a patch-clamp setup, you place neurons or a slice of brain tissue under a microscope, and you can actually visualize the neuron, and you use micromanipulators, very fine manipulators, to move this glass pipette or electrode to the neuron, and inside the electrode is actually a solution which tries to approximate the internal solution inside a neuron. So the same chemicals and concentration of ions that occur inside the neuron, we put those in the pipette.
And then you approach with a micromanipulator up to the neuron and touch this glass electrode up against the surface of the neuron. And then you apply slight suction to the electrode, and you break a hole into the cell, and this makes a continuous solution between the pipette or the electrode and the internal solution of the cell, and in this way, you can record the electrical currents in the cell.
What other new technologies will be important?
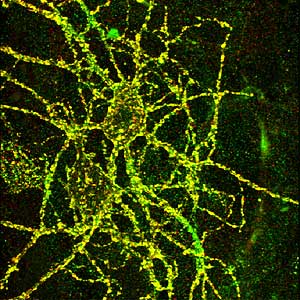
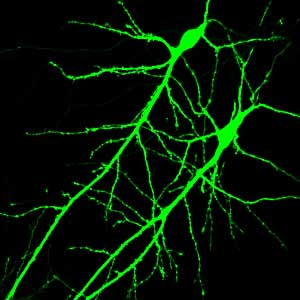
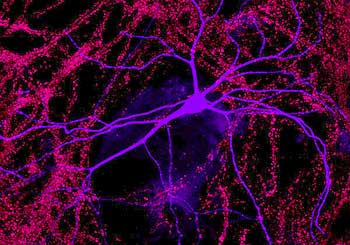
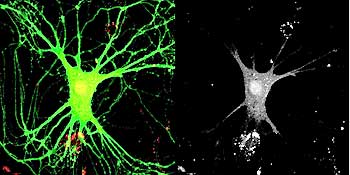
Imagining technologies have really incredibly improved over the last decade, and one of the more powerful techniques we use today is called “live cell imaging,” or “time-lapse imaging,” and in this case we can actually visually see the receptors in the cell. And the way you do this is we take advantage of jellyfish, which have evolved to make proteins that fluoresce-they actually give off light. And so what we do is attach this jellyfish protein to our receptors and then express this modified receptor in neurons. And what happens is the neurons make the receptor as normal, but it turns out that this receptor now glows green, so you can actually visually see the receptor as it’s made in the cell and exported out into the dendrite and incorporated into the synapse.
So we can actually watch that over real-time as the receptor moves through the cell and goes to the synapse, and-as I mentioned earlier-receptors can be endocytosed, so we can actually watch the receptors be removed from the synapse and taken back into the cell. So it’s a way of really looking at the dynamic properties of these receptors in the neuron, and this is one way we found out how dynamic these receptors are. For a long time, we thought the receptors were pretty static, that they would just stay in the synapse for days, but now we know that they are constantly moving. They’re moving in and out of the synapses on a minute time scale.
How would the brain slice procedure be used to see this?
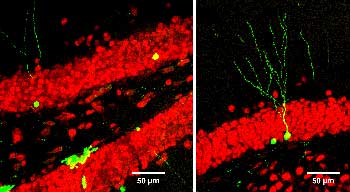
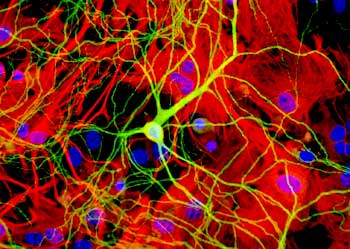
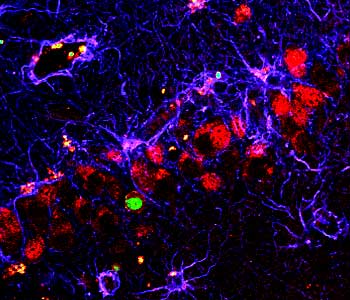
A couple of the major preparations we use are brain slices and cultures of brain neurons. And so, to really study the network properties of the brain, the natural connections in the brain, we want to use a brain slice, so we take a section of brain and maintain all the normal connections between neurons, so we can record network activity in those slices.
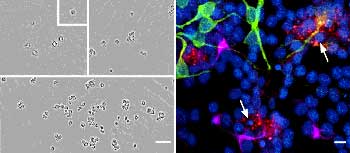
For other reasons, we want to actually study a neuron in isolation, and in that case, we take the brain and we dissociate the cells, and we grow them in a dish under conditions where there are relatively few connections, and in that case it’s much easier to image them, to see them, because they’re in isolation. So in many cases, we do some of the live imaging with these fluorescent proteins in the dissociated cell culture.
In contrast, we do a lot of the electrophysiology, the studies of long-term potentiation and long-term depression, in slices, where we have maintained the natural connections between neurons.
What are the major questions that motivate you?
One of the major questions that remain now that we really understand synaptic transmission at a molecular level is to understand these long-lasting changes. We understand some of the basic principles of how you change synaptic transmission, how it can be plastic, how you can modify receptor structure or the release of neurotransmitters to potentiate a synapse. But how do you maintain that potentiated state for days or weeks or years? We know that the receptors, the proteins themselves that are modified, only last a few days, so they will actually turnover–that is, they will be broken down, degraded, and new ones will replace those receptors. But somehow that synapse is still potentiated, so in spite of the fact that the receptors have been totally turned over and replaced, that synapse is still potentiated. So there must be some self-perpetuation process at work.
So a major question remains: how are these synapses potentiated for such long periods in spite of the fact that the receptors are turning over? So if we can understand, for example, how these processes of increased insertion of receptors, and increased phosphorylation of the receptors, are perpetuated over periods of weeks or months in spite of the fact that receptors are turning over constantly at the synapses.
What do you most want teachers to know?
The more we understand about how synapses work and how they’re modified, the more we’ll understand how the brain works. And of course, this relates to many processes in the brain, including disease states. Many neurological diseases are breakdowns in these systems, and it’s clear that glutamate receptors are involved in many diseases.
Stroke is a classic example, where glutamate receptors are key in the neuronal damage that occurs during the stroke; epilepsy, and a variety of other diseases–Lou Gehrig’s Disease, ALS. Glutamate receptors play a key role in those diseases. So I think if we can understand how to regulate receptors, we’re going to also affect and have better insight into diseases.
Of course memory is a problem for all of us, especially as we get older, so if we really understand how memory and learning occurs at a molecular level, we’ll be able to possibly intervene, and I think hopefully within my lifetime there will be ways to intervene, and to actually develop what we call a cognitive enhancers–things that will enhance memories, and for me, of course, that means enhance synaptic connections in the brain.
What are the bioethical considerations involved?
All of these studies use animal models, and this is really key for our understanding of brain function. If we could study bacteria or yeast cells, that would actually make our life easier, because they’re much easier organisms to study. But of course yeast and bacteria do not have nervous systems, so we really rely on animal systems to study brain function, and I think these types of studies have played a large role in the development of therapeutics for many neurological diseases, and without animal research, this would not have occurred.
Are there alternate ways available?
One problem with neuronal tissue and neurons, in general, is that it’s very hard to develop cell lines of neurons, and so that’s one thing that’s hindered the field, and so we have to use what we call primary cultures, cells that come directly from animals. So we would love to be able to have cell lines that we could use, and just thaw them out and grow them up for our experiments, but so far that’s usually not been the case in studying the ‘naturally occurring’ complement of molecules that are expressed in neurons, although cell lines are often used to study particular molecules-channels, for example-in isolation.