








Rediscovering Biology: Molecular to Global Perspectives
Evolution and Phylogenetics Expert Interview Transcripts: Carl R. Woese, PhD
 Professor of Microbiology
Professor of Microbiology
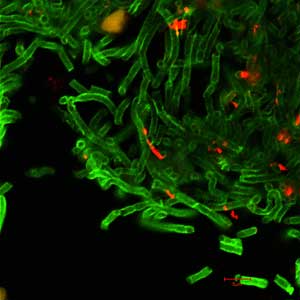
Professor Woese of the University of Illinois, is a self-described molecular biologist turned evolutionist. Woese used ribosomal RNA as an evolutionary record to identify archaea as a huge and diverse group, separate from the prokaryotes (bacteria) and the eukaryotes. As a result, the entire “Tree of Life” as we know it was re-drawn into these three domains.
Interview Transcript
Professor Woese of the University of Illinois, is a self-described molecular biologist turned evolutionist. Woese used ribosomal RNA as an evolutionary record to identify archaea as a huge and diverse group, separate from the prokaryotes (bacteria) and the eukaryotes. As a result, the entire “Tree of Life” as we know it was re-drawn into these three domains.
Can you define molecular systematics?
Molecular systematics involves viewing the organism from the molecular level-in other words, nowadays, in terms of sequences of micro molecules, the proteins and the nucleic acids. On that level, one can see things that are not seen when you look more grossly, say under the microscope or with the naked eye. It has been said that the molecular level is the most delicate expression of the phenotype of an organism. The molecular systematist views the organism from the level of the molecule. The classical systematists were forced to work with the level of the whole organism or looking under the microscope at individual cells. There are many secrets hidden there that you cannot see at the higher levels-looking under the microscope or looking at the whole organism per se.
What is ribosomal RNA?
The ribosome is the main machine in the cell. It is the molecular mechanism that takes the information stored in the gene and translates it into information in protein sequence. It’s just like translating a tape written in one language into a tape written in another. And ribosomal RNA is the core of the ribosome. The ribosome comprises both RNA and protein and in effect the ribosome comprises also two subunits-one is about double the size of the other-each of which contains a core of RNA surrounded by special proteins.
Define archaea?
The archaea are what has been called at various times the third form of life. What is meant by that is the following and it has to be set in a historical context. For years and years, people thought that at base there were two types of organisms on this planet. The so-called eukaryotes are represented by animals, plants, and everything you can see with the naked eye that is living, including certain microscopic things, such as the familiar paramecium. And then there were the bacteria.
Now the telling difference between the two at the level of the cell itself, i.e., looking under the microscope and into the interior structure of the cell, is that the eukaryotic cell contains a membrane-bounded nucleus, so that it sticks out. It’s one of the first things you’d see looking at a cell under the microscope. Whereas in the case of the bacteria, the other form of life it is the case that there, they have no nuclear membrane containing the genes, as the eukaryotes do. Nevertheless, they have genes. It’s just that they’re not enclosed in a bag.
Could you define horizontal gene transfer?
Horizontal gene transfer is a mechanism by which an alien gene from a source “organism X,” moves into, or is taken into, a given cell and is somehow incorporated into the genetic complement of the cell. That gene had previously not been in the cell, although in certain cases something very like it might have been. The best examples you have before you today, of course, involve antibiotic resistance, and I think just about every reading adult is familiar with antibiotic resistance. And this occurs usually when a cell that is killed by the antibiotic such as a staphylococcus or a streptococcus, when that cell picks up a package of genes called a plasmid, among those genes might be one that produces a protein product that enables the organism to somehow avoid, inactivate, overcome the antibiotic.
And this has to do with all the furor we have about which antibiotics can be used in which medical conditions. With certain antibiotics, the resistance genes for them are not very prevalent in the environment, so that we save those for the desperate cases where all the other antibiotics don’t work.
Can you define extreme thermopiles?
Extreme thermopiles are a subset of the extremophiles. Extremophiles are, by definition, organisms that grow under what we consider extreme environments: extremes of temperature, extremes of salinity, extremes of acidity or alkalinity — that sort of thing. And then of course extreme thermophiles are those that grow at extremes of temperature. There are some known that will grow up above the normal boiling point of water, but not by a lot — maybe 10 or 20 degrees above it.
What is the role of molecular data in discerning evolutionary relationships?Molecular data have made an absolutely profound revolution in the way the evolutionist can look at the world. As I said previously, molecular sequences are an extremely delicate expression of the organism’s phenotype. They are more subtle and more informative than any other characteristic the cell has-the microscopic characteristics.
Without molecular data, most of the living world would not be understandable in taxonomic terms. By this, I mean that when classical biologists defined the categories of organisms they worked essentially with animals and plants. Now animals and plants are actually only a small fraction of the diversity of life on this planet. Most of that diversity lies in the bacterial world.
It isn’t that microbiologists didn’t want to understand the evolutionary relationships, the natural relationships among bacteria. It’s that, given the classical criteria, the shapes of the cell, and the physiological reactions, they were unable to infer from these qualities the relationships among organisms.
But, the minute the molecular data came around it was obvious that relationships among bacteria from the most distant to the closest could all be sorted out and bacteria could be classified just as well as any other of the living systems, animals, and plants for example. So it has had a great unifying effect on our understanding of how all life on earth is related to all other life on earth.
How has that changed our view of those relationships?
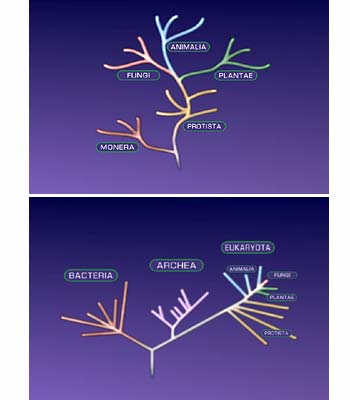
Well, the principal one that came sort of as a shock initially was that for a long time biologists had assumed that, as I said, that there are two types of life on this planet, at base-two primary types. One being the eukaryotes characterized by cells with a membrane-bounded nuclei. And the other was the prokaryotes, characterized by cells that did not have membranes around their genes in the nucleus. Now, they made the mistake of assuming that those so-called prokaryotes were all related to one another because of this. In other words, because they’re not eukaryotes they’re all the same kind of thing. They’re all kin. That was a bad mistake.
Now the minute you got down to the molecular level and began defining relationships in terms of similarities of molecular sequences you immediately saw that the so-called monolithic grouping prokaryotes was not that. It actually comprised two distinct groupings, neither of which was more related to the other than either of them was related to the eukaryotes.
How did they come to be identified?
This is a story that goes way back. As a young Ph.D., I developed an interest in evolutionary problems-one in particular-and that was how the translation apparatus, the central machine of the cell, came to be. This is such a remarkable apparatus in that it is so complex. It has to have an interesting story behind it-how it came to be. So it also became obvious that the only way you could get at this was to do your studies on the molecular level. Now you’re asking the question: How did translation evolve? It represents a level of evolutionary questions that Darwin couldn’t possibly even formulate because there was no knowledge of the molecular world at that point. So these are what I like to call “pre-Darwinian questions.” They go deeper into the evolutionary course than a classical evolutionist could go.
Now it’s clear that in order to understand anything about the evolution of various components in the cell — the pre-Darwinian types of evolution — one has to have a tree of relationships among the organisms, all the bacteria.
And when I began my work, the only relationships that were known were those inferred by the classical evolutionists which were indeed the phylogeny, the tree, of the plants, the phylogeny of the animals. The whole microbial world was terra incognito.
So the first thing to do if I’m going to study evolution of translation is to fix the situation and create a tree that encompasses all of life, and it was clear at the time that this probably could be done by comparing the sequences of particular molecules, the one that was obviously the most useful because it was the easiest to handle in the laboratory was the one called ribosomal RNA.
And so began an extensive survey of the sequences of ribosomal RNAs. And at that time, one could-I’m talking here “1970ish”-one could not sequence the molecule outright but one could cut it into pieces, separate the pieces, and then characterize each of the pieces. That’s producing a catalog of fragments that were characteristic of the ribosomal RNA of a given organism. And this is the way we began and we accumulated catalogs from a great many bacteria, up to 400 or so and these catalogs, when compared, would allow us to tell how close the various bacteria were to one another in ancestry.
The more similar the catalog of pieces the more recently the organisms would have shared a common ancestor. And from this, you could then build trees through the usual tree-producing algorithms.
There are two main things that you can use this framework for, and by this, I mean things that are to some extent, at least, outside the realm of science. Mankind has always asked the question: Where did we come from? And Darwin began to provide the answer to that in saying we all evolved from a common ancestor.
Humans had a primate ancestor. The primates had a mammalian ancestor; that mammal had a vertebrate ancestor, and so on down the line. But this is only talking in terms of animals. To really get deep into this question, where we came from, you need to talk about how all life is related one to the other, all living systems, and that can only be done by inferring the relationships from molecular data.
Now the second thing that the universal phylogenic tree gives you, placing organisms on it as you find them, is that it allows you to become much more knowledgeable about what’s going on in the environment. I mean the question of the nature of the biosphere is not only a deep biological question, but it is also critical from a practical point of view, since man is putting many, many things into the biosphere in great amounts that have the potential for changing the nature of the biosphere. We don’t know whether they will or not, but if you put hundreds of tons of something into the atmosphere per year, you can bet that changes will somehow occur. It’s not just a minor perturbation.
What changes occurred to the tree after your study of ribosomal RNA?
The first thing, the most superficial one, is that life actually comprised three major types of organisms, not two: eukaryotes, bacteria, and archaea; not just eukaryotes and the sum of bacteria and archaea prokaryotes. But if you know very little you only make very general conjectures. If you know more, your conjectures may be a bit more specific. So it’s been a sort of a case of turning up the light in a dark room. You begin to see more and more as the light intensity increases. So I’ve gone from a state of sort of just general speculation to one in which I can now talk in somewhat specific terms about what might have happened way back four billion, three and a half billion years ago.
How would you define evolution theory?
One of the things that one expects of a theory is that it makes predictions that are testable. Now, this is not a necessary property of a theory but it’s better if it makes predictions that are testable. If it serves only an explanatory role and it can do so in a fairly detailed way, that’s still very good. And if it has an effect upon the general way we think about things if it turns our heads in one direction rather than any of the other directions, and that’s a productive direction, that’s good. So these are sort of the three functions of theory. One is an explanation, one is prediction, and the other is its conceptual impact.
Now clearly one can’t do evolutionary experiments in the sense of evolving animals over again. So there’s very little that an evolutionary theory can do in the way of making testable predictions. Nevertheless, what it can do is lead you to analyze the data that are available in a sense of the experiments that nature itself has done in such a way that you can then create a consistent picture that grows more detailed and more consistent as it goes along.
And that is in a sense what scientists take as the mark of something that’s “true;” that the more new facts one brings into play, the sharper the picture becomes. It doesn’t tend to fall apart. It tends to refine itself. And while this is not using experimental tests to test the theory, evolutionary theory does give you-especially now with the capacity to sequence genomes-it gives you a very detailed consistent picture.
I think that one cannot understand biology except in an evolutionary concept. It’s as simple as that. Biology falls apart if you take away evolution. You have just this odd collection of things, very different, very complex, and you don’t really know how they relate to one another. It does really no good scientifically to say they’re all placed here separately. Then where do you go from there? How does that help you understand life? It doesn’t. Whereas evolution is a unifying force in our understanding and it does a good job of it.
Is rRNA easy to handle in lab?
Ribosomal RNA is universal. It occurs in every living organism. In a typical bacterial cell there are on the order of ten or more thousands of copies of the ribosome. So that this is a piece of nucleic acid that is ten thousand fold amplified over any other particular gene, which is expressed in only one or two copies. So for that reason you have the amount of it running in your favor and the laboratory techniques for extracting ribosomes had become very sophisticated by the time I started. It’s very easy to break open a cell and centrifuge out the ribosomes, for example, and then separate the large subunit of the ribosome from the small, and then extract the RNA from the small subunit, in this case.
These were routine operations. Whereas, for example, to isolate any other given protein, that would have been a rather lengthy thing to do, and many of them couldn’t even be isolated at that point. But when of course their genes could be sequenced, which was later, then more molecules could be brought to play in this game of doing genealogies. Their sequences could be determined through the sequencing of their genes.
But to begin with it had to be something like ribosomal RNA. It was the only feasible protein to isolate. It was large, ubiquitous, and easy to isolate.
What techniques did you use?
We were not able to look at the sequence of ribosomal RNA as a whole. We had to break it into fragments of pieces from a stretch of 5 nucleotides up to maybe 20 nucleotides. It’s like taking a play and breaking it into words or phrases, but you still can know something about the play by the frequency of word use and that sort of thing.
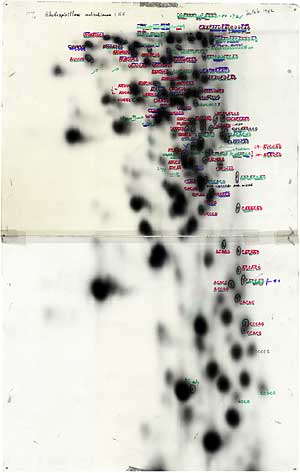
So what we did at that point was to grow the cells that we were about to extract the ribosomal RNA from. We grew those cells in a P32 radioactively labeled phosphate medium. And then of course, when the ribosomal RNA was isolated, since the backbone of a nuclear acid chain is phosphate molecules, this thing was highly labeled. So you could expose it-very small amounts of it-to a photographic film and see a darkening of the film.
So if you had some sort of procedure that would separate out these pieces of the ribosomal RNA, produced enzymatically, from one another say by electrophoresis-actually in our case it was two-dimensional paper electrophoresis-you could then expose that paper to film and then see what you’ve got. And what we got was a series of spots in a particular kind of pattern, and that showed you where on the paper the pieces of ribosomal RNA were located. And then you had to do further analysis of a similar kind to determine the exact sequence of each of the so-called oligonucleotides that were represented by a given spot on the radio audiogram.
So how has that data now been used?
You compare the sequences of a given ribosomal RNA from one organism to the next and then to the next and the next. And, crudely speaking, the closeness of these sequences to one another, their degree of similarity, determines when–relatively speaking–they had a recent common ancestor. And that so-called phylogenetic analysis is sort of the backbone of how a molecular evolutionist approaches genomes.
And then there’s also another question of what genes are there in the genome. This is a question that could never be answered before genome sequencing. If you didn’t find a particular function manifested, say in a bacterium, some sort of capacity to metabolize this or that sugar, then you weren’t absolutely sure the gene was there. The gene could have been there and making the protein. The gene could have been there but inactive for some reason, not being expressed. It could have been there but inactive for the reason of having mutations in it that rendered the expression product, the protein nonfunctional.
But nowadays with full genome sequencing, you can say whether a gene or anything like it is there. So this question of how you interpret a negative finding has been placed in a new context. It’s much more informative.
What are some of the complications of full genome sequencing?
Well, back in the day, which is even up to the early ’90s, back in the times when one had only a few molecules to play with, to take up your attention, the picture of a relationships-the universal tree-looked very simple and nice from the from a molecular perspective. There it was. And you could see this group branching first from the root base of the tree and then a little later another two groups spawning out–the first one being the bacteria and the other two being the archaea and the eukaryotes. It seemed like everything was falling in place.
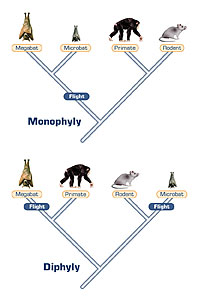
However, up popped the specter of horizontal gene transfer in a big way, because when one attempted to infer the universal tree structure not from ribosomal RNA sequences but from the sequences of various proteins, you tended to get fundamentally different branching orders because Protein X may give you one ordering, Protein Y may give you a somewhat different ordering, Protein Z an entirely different ordering and on and on.
Now if the proteins had given a different ordering that all agreed with one another, one protein to the next, then they would have said something is wrong with the ribosomal RNA inferred universal tree but that’s not what happened. Most of the proteins’ trees disagreed with the branching of the ribosomal tree, but they also disagreed with one another.
So the only simple way this could be explained was to say that somewhere along the line these things have been influenced by the importation, the horizontal transfer, of genes that don’t really bespeak the lineage of the organism. They came in at a certain point and made the lineage appear to change. But it was only the gene bringing its own history with it, not the history of the organism. So this served as confusion, as noise thrown into the message.
And as things have now shaken out, it’s clear that the ribosomal RNA-inferred tree is undoubtedly the correct one because there are indeed certain proteins that agree with it, whereas the remaining proteins that don’t agree with it also don’t agree with one another, so they aren’t really telling you anything.
And the stability of the ribosomal RNA genes in the evolutionary sense, they are not readily subject to horizontal gene transfer, so that they tend very highly to remain fixed with the organism. Therefore, they bespeak the organism’s lineage as a genealogy. They’re the most accurate and reliable sources of information on the genealogy that we have. If you’re going to try to understand how translation evolved, you should be working with a molecule that’s part of the system.
Can you describe a “wow moment”?
Well, it was when the archaea were first discovered, because I, along with every other biologist in the world, as far as I know, had made this false assumption that there were two types of cells, two types of organisms on this planet: the so-called prokaryotes and the eukaryotes. So we had gotten a pretty good read, a pretty good signature, that said, okay, if the ribosomal RNA gives this kind of catalog of all of the nucleotides then it’s a prokaryote. If it gives that kind of catalog then it’s a eukaryote.
I can still picture very clearly in my mind standing at the so-called light board where we used to analyze these films. You had light transmitted through the film so you could see the dark spots. I remember saying, well, in this particular archaea, the nucleotide characteristic of the prokaryotes is not here, and neither are these two.
So I eagerly continued to infer that sequences of these things by the techniques we had, and write them on the master film again next to each spot. And by the time I finished it was very, very clear that this was something of a different kind. It was neither the so-called prokaryotic kind of cell, nor of the eukaryotic kind.
And so it wasn’t long before I realized, “oh my God, we’ve made a mistake here in assuming that all prokaryotes were the same kind.” Because what prokaryote was by definition was a little tiny bacterial looking thing. But this just didn’t fit with its signature. It didn’t have a signature because we were only looking at the first one. It just didn’t fit the signature of the bacteria or the signature of the eukaryotes.
So that was really a thrilling moment. And of course right away there was a rush, one, to repeat the experiment on the assumption that somewhere something might have gone wrong somewhere. Well, it hadn’t. So then we took the relatives of the organism we had looked at, and then looked at them and saw that they all don’t have the bacterial signature, and they all don’t have the eukaryotic signature, but they do now begin to define a signature of their own. That was thrilling. But it’s also thrilling to sit there and look at the universal tree for the first time. This is it. This is the complete picture.
What are the shortcomings of present theory?
Well, we’re on solid ground enough that when things vex you, when they don’t seem to fit, you could be almost certain that you’re gonna find out something new. It’s not that what you do know is gonna fall apart. And that’s been sort of the case again and again.
For example, we’re going to have to get used to the idea that the universal tree is not a normal tree. Okay? A tree is after all only a representation of what you know about the relationships among organisms. There was a time apparently before even one reaches the root of the universal tree when the organisms that were evolving were in very intimate contact with one another via horizontal gene transfer. In fact, they helped one another bootstrap into becoming modern cells.
Now, at that time cells were of a nature where nothing was permanent. Genes were shuffling in and out-at least on an evolutionary time scale. So if you have a genealogical trace started, it could easily have been (and sometimes was) over sufficient time just erased by horizontal gene transfer.