







Rediscovering Biology: Molecular to Global Perspectives
Genetics of Development Expert Interview Transcript: Judith Eisen, Ph.D.
 Professor of Biology
Professor of Biology
Eisen is a professor at the Department of Biology at the University of Oregon. She studies how neuronal diversity is generated during development, including how the correct number of cells are specified for specific neuronal fates at particular times and in particular locations. She specifically works on the patterning of neurons and neural crest cells in embryonic zebrafish.
Interview Transcript
Interview with Judith Eisen, Ph.D. Eisen is a professor at the Department of Biology at the University of Oregon. She studies how neuronal diversity is generated during development, including how the correct number of cells are specified for specific neuronal fates at particular times and in particular locations. She specifically works on the patterning of neurons and neural crest cells in embryonic zebrafish.
Can you provide a brief history your research and why you chose to study zebrafish?
The history of zebrafish use really started here at the University of Oregon with George Streisinger who was the first person to try to use zebrafish for a genetic analysis of vertebrate development. George came from what you might call the original school of molecular biology. When he came here in the late sixties, he brought with him a vision of trying to do a genetic dissection of vertebrate development, and he needed to choose an experimental organism to do that. There were several other people that were trying to do similar things. Sydney Brenner was trying to do this kind of thing with a small soil nematode called C. elegans and Seymour Benzer was trying to do similar things with the fruit fly, Drosophila.
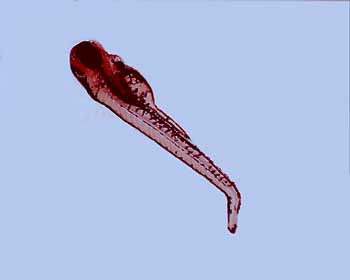
Nobody else was trying to do this with such a complicated animal as a vertebrate. So Streisinger needed to search around and find an animal to use and he chose zebrafish. Zebrafish have a lot of advantages for doing this kind of work. They’re easy to raise in a laboratory setting; they breed very readily-a single zebrafish adult female can give up to several hundred eggs on a weekly basis. Having large numbers of eggs accessible is very important for doing a genetic analysis. The eggs are laid into the water column and fertilized by the males in the water column, so all of development takes place outside of the mother and that’s very important. It means that every stage is accessible and can be studied in a lot of detail. That’s in contrast to another organism like the mouse, which is a very popular organism for doing these kinds of studies. Of course in a mouse, all of development takes place inside of the mother and that makes it less accessible-more difficult to study.
Zebrafish embryos have a number of other kinds of advantages also. The embryos are optically clear, which means that by putting them under a microscope and using appropriate kinds of optics, we can actually image every cell in the embryo. We can watch as its development unfolds, as the embryo goes from one cell to two to four to eight to sixteen. Then at even later stages, we can image individual muscle cells, individual nerve cells, and other individual cells and actually watch and see what they do. That’s something that’s extraordinarily difficult to do in other vertebrate organisms, although it can be done in C. elegans and in fruit flies. Of course, those are both model organisms for doing these kinds of studies, but they’re not vertebrates. If you really want to understand how a vertebrate develops, you need to use a vertebrate organism. Although we can learn many, many principles from studying these other organisms.
This was George’s vision when he came here and started his lab in the early ’70s and started working on zebrafish. Then about the same time, Chuck Kimmel-who was already here-started using zebrafish to do an analysis of nervous system development. Eventually, there were a number of laboratories here that were all working on zebrafish, trying to use the fish to understand the genetic mechanisms, the molecular mechanisms, and the cellular mechanisms underlying vertebrate development. So the fish facility is shared by all of these laboratories.
Why is it important to study vertebrates as opposed to the fly and the worm?
Over the last several decades, we’ve learned an enormous amount about how embryonic development happens by studying C. elegans and by studying Drosophila melanogaster. One of the probably most amazing and most interesting things that we’ve learned is that a lot of the genes that govern development in these organisms also govern development in vertebrate organisms. By studying these organisms, we can learn an enormous amount. But they’re not vertebrate. If you think about the body plan of a human and the body plan of something like a fruit fly or a worm, we’re obviously very different. Even if the same genes and the same mechanisms are regulating that, how we turn out is completely different. So to really understand how a vertebrate turns out the way it does, you have to look at a vertebrate. You can’t learn those precise things from looking at something that’s not a vertebrate.
Can you go into more detail about the zebrafish embryos and how they are used in genetic research?
One of the really great advantages of zebrafish embryos is that they’re really accessible for experimental embryology sorts of approaches. That means that we can move cells from one embryo to another embryo and learn how those cells respond. We can move cells at different times, or from embryos of one genotype to embryos of another genotype and understand how that influences the development of those cells. We can do this not just with clumps of cells, but we can actually move a single cell between embryos and do it on a regular basis.
So that’s something that is different or impossible on any other model organisms. We can also very easily inject things into zebrafish embryos. We can inject messenger RNAs; we can inject things that block the function of a gene; we can inject dyes of various sorts into single cells at various different stages. So this gives us a whole host of techniques to use that are embryological, but also allows us to combine those with genetics so that we can do these kinds of experiments in embryos that have different mutations-they have different genotypes-and it really allows us to gather an enormous amount of information.
Can you talk about your research specifically?
One of the things that we’re really interested in studying in my laboratory is how different kinds of neurons are formed. There are lots of different kinds of neurons. There are motor neurons that enervate muscles, sensory neurons that take in information from the outside world, and inter-neurons that connect the two. We’re particularly interested in understanding how motor neurons are formed and how they then grow out to the right part of the body and to the correct muscle.
If you think about our bodies, this is really important. We have lots and lots of different muscles and different parts of our bodies, and each kind of muscle is enervated by a particular kind of motor neuron. If the wrong motor neuron went to the wrong place, you wouldn’t be able to walk or move around in the ways that we do. So it’s a very challenging sort of question.
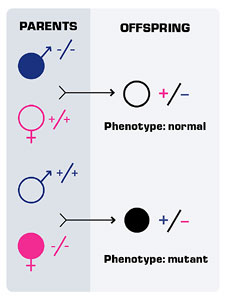
One of the things that we know from work that’s been done in other species-particularly in chickens and mice-is that a particular protein, called Sonic Hedgehog protein, is very important for determining whether particular nerve cells are going to be motor neurons or not. A few years ago, a mutation in the Sonic Hedgehog gene was isolated in zebrafish. The surprising thing was that mutants for the Sonic Hedgehog gene in zebrafish still had motor neurons. This is a big contrast to mouse mutants in the Sonic Hedgehog gene, which don’t have motor neurons. So we wondered what was going on, and of course, there are several possibilities. One possibility is that things are different in zebrafish than they are in the mouse. That would be a very disheartening possibility, because we want to be able to use zebrafish and mice and other organisms as models: if one or more of them really is not going to serve as a model, that would be a real problem for the kind of research that we’re doing. But there are other possibilities too. Another possibility that we thought was very likely was that there might be additional Hedgehog proteins in zebrafish.
We know from the work of a number of other laboratories that this is the case — that there are additional Hedgehog genes in zebrafish. These genes encode molecules that are related to Sonic Hedgehog, but they’re not identical. We also know from these other laboratories that two of these proteins, one called Echidna Hedgehog and one called Tiggywinkle Hedgehog are expressed in the embryo at the right place and the right time to be working together with Sonic Hedgehog during the formation of motor neurons. We reasoned that maybe if we got rid of all of the Hedgehog proteins that we would find that there were no more motor neurons. So Kate Lewis, who’s a postdoctoral fellow in my laboratory, actually did those experiments. She got rid of all of the Hedgehog proteins and this turned out to be a difficult set of experiments to do. I said that we have a mutation in the Sonic Hedgehog gene, and that means that embryos that have that mutation don’t make Sonic Hedgehog protein. But we don’t have mutations in the Echidna Hedgehog gene or the Tiggywinkle Hedgehog gene, and when Kate began these experiments, we didn’t have a functional way of knocking out those proteins.
What Kate did was to make embryos that have mutations in structures that produce either Tiggywinkle Hedgehog or Echidna Hedgehog. The reasoning for this was that if you delete the structure that produces the protein, then you don’t have the protein.
To do this, she had to have embryos that carry three different mutations. A mutation in the Sonic Hedgehog gene, a mutation in a structure that produces Echidna Hedgehog, and a mutation in a structure that produces Tiggywinkle Hedgehog. What she found is in those mutant embryos –embryos carrying all three of those mutations-the motor neurons were essentially gone. She’s repeated those experiments now, using combinations of morpholinos injected into embryos. Mostly what she’s done is inject morpholinos for Tiggywinkle Hedgehog and for Echidna Hedgehog into embryos that are carrying the mutation in the Sonic Hedgehog gene. These embryos don’t make Sonic Hedgehog and she’s knocked out the ability of them to translate the proteins for Echidna Hedgehog and for Tiggywinkle Hedgehog. So just like the triple mutants, these embryos don’t have any Hedgehog protein, and again, the motor neurons are essentially all gone.
If you take away only one of the Hedgehog proteins and leave two, you reduce the number of motor neurons, so it’s intermediate in between the levels that you normally have, and the levels that you have when you get rid of all of the Hedgehogs. If you just get rid of one of them, though, you hardly see any disruption in the number of motor neurons. So that means that these Hedgehog proteins are actually working together during the formation of motor neurons. And any one of the Hedgehog proteins is dispensable, but you need to have at least two of them.
So the presence of the hedgehog protein is what can be referred to as the gradient?
Right. One of the things that is really interesting is that Hedgehog proteins, and particularly Sonic Hedgehog protein in other animals, has been shown to act in a gradient fashion. What that means is, different levels of Sonic Hedgehog protein cause formation of different types of cells. That’s something we’re also really interested in understanding in the zebrafish. How does that work?
One way to do this would be to measure the levels of Hedgehog protein. It turns out that that’s a very difficult thing to do, so what we do instead is look at things that are downstream of the signaling process. So something that happens after the signaling process and provides us a readout or an assay to tell us that the signaling process has occurred.
What we mostly look at are the genes that cells express and what those cells turn into. We know that some of those genes directly respond to Hedgehog and that different levels of Hedgehog are required to turn them on. If we see expression of a gene that requires a lot of Hedgehog to turn it on, we would expect to see that very close to the source of Hedgehog, and that’s what we see. But in contrast, a gene that’s turned on by a much lower amount of Hedgehog, we’d expect to see further away from the source, and once again, that’s what we see. That gives us a readout that the levels of Hedgehog protein start in one particular place, and they move away from that place, and as they do, the concentration decreases. This is a concentration gradient-dependent effect, or sometimes we call it a “morphogen.” A morphogen is a molecule that has different effects at different concentration levels.
Is the research that you’re doing referred to as “cell-staining”?
In order to see where cells are expressing particular genes, we have to have a readout. Our readout is to either look for the messenger-RNA that’s produced when a gene is turned on, or to look for the protein that’s produced from that mRNA. And to do this, we do what we call “staining the cells.” What we’re doing is using a reagent that actually recognizes the mRNA itself, and it gives us a colored reaction product, so we can see that cells are blue if they have started to make this mRNA. Or we’re using a reagent that allows us to recognize the protein — that’s called an antibody — so that we can see that cells are yellow or red or some other color if they’ve started to make this particular protein.
So the motor neurons that we study express some very specific genes. One gene is called Eyelet 2, and then other motor neurons express a related gene called Eyelet 1. We can use these as a readout to see whether they have received Sonic Hedgehog signaling, or signaling from the other Hedgehog proteins. We can stain the embryos for the presence of mRNA — either for Eyelet 1, or for Eyelet 2, or for both of them — and we can see which cells are expressing those genes. Because we know from our experiments in which we took Hedgehog away that those cells don’t form in the absence of Hedgehog, then it tells us if we see those genes expressed in cells at the right place, at the right time, that Sonic Hedgehog signaling-or Hedgehog signaling in general-has to have occurred. We can also use an antibody that stains the Eyelet 1 and Eyelet 2 proteins and so that lets us see that the protein is made, not just the messenger-RNA.
Can you define in-situ hybridization?
In-situ hybridization is a process that allows us to see which cells are expressing a particular mRNA. The way we do that is we make a probe that is similar to that mRNA, but it’s an anti-sense probe. That is, the sequence of nucleotides are the ones that bind to the original mRNA. Then we tag that anti-sense probe, so we can then react it with something that will reveal color to us in the cells that are expressing that mRNA.
What are the big unanswered questions that you have in your research?
There are a lot of big unanswered questions in trying to understand developmental biology. One of the questions that I’m really excited about trying to understand is what is it that makes different nervous systems different from one another. If you think about something like a fruit fly, and something like a human, or a zebrafish or a mouse, the nervous system architecture is so different, and yet the same sets of genes are probably involved in the early development of those nervous systems. So a zebrafish and a mouse and a human are pretty similar, but really different in a fruit fly, and I would really like to understand how it is that those differences in architecture are established, and how the genes are deployed in different ways, at different times, in different cells to make things be so different in those two different types of nervous systems. And what that means in an evolutionary sense. Because at some point, we and fruit flies, and zebrafish descended from a common ancestor. So what was the nervous system of that ancestor like?
We can take a specific gene out of a zebrafish and we can put that into a fruit fly that has a mutation so it’s own copy of that gene doesn’t work, and under some circumstances, the gene will substitute perfectly well. Now, what does it do? Does it make zebrafish structures in that fruit fly? No. It still tells the fruit fly to make the fruit fly structures that that gene would normally regulate. So that means that the zebrafish gene and the fruit fly gene are actually close enough in structure that they can actually carry out the same function. That’s not true for every gene. Some genes that have been tested are very diverged between a fruit fly and a zebrafish and the zebrafish gene simply doesn’t work in the fruit fly or it doesn’t work correctly, but in some cases it works perfectly well.
Can you talk more about cell-cell signaling?
The process of development can be characterized a series of signaling interactions between cells. A cell that is starting to do something can be signaled by another cell, and that will change what it’s starting to do and send it off along a different developmental pathway. What happens during early embryonic development is cells rearrange-they get new neighbors, and that provides them with new signals that they haven’t experienced before, or they haven’t experienced in the same way, and that sets them off on different paths of development.
Within the context of the cells that we’re studying-motor neurons-we know that there are lots of cells that could become motor neurons, but only some of them do. They do because of signaling interactions that they undergo, but once the cell has made the decision to become a motor neuron, and turned on the appropriate genes, it signals to all the cells surrounding it and tells them, “don’t do that-do something else.” If we take that signal away, then all of the surrounding cells will become motor neurons as well. We also know that there are different signals that impinge on these cells at different times that are important for their individual identities. We know that because we can pick a cell up from one position and put it into a different position, and if we do that late in its development, it develops in a way that’s appropriate for the place that we took it from. It doesn’t change what it does. But if we do it early in development, it changes. It now becomes a different kind of motor neuron that’s appropriate for its new place in the embryo.
What that tells us is there has to be some signal that the cell responds to at an early stage in its development and that that signal is really critical for telling the cell what kind of motor neuron to become. But at later stages the cell is not responsive to that signal anymore, and so the cell is undergoing some sort of change. Now, we of course know nothing about what those signals are. There may be one, there could be many of them-we don’t know that either, but that’s a question that we’re really, really excited about addressing in my lab.
A lot of our research is aimed at trying to understand the nature of those signals-what cells are producing the signals, what molecules constitute the signals, how many there are, when they’re produced and how the cells are responding to them so that this changes the course of their development.