







Rediscovering Biology: Molecular to Global Perspectives
Genetics of Development Expert Interview Transcript: John Postlethwait, Ph.D.
 Professor of Biology
Professor of Biology
Postlethwait is a professor of biology at the University of Oregon, Institute of Neuroscience. He studies the genetic regulation of animal development including development of the nervous system, the mechanisms of sex determination, the origin of novel morphologies in evolution and the evolution of the vertebrate genome.
Interview Transcript
Interview with John Postlethwait, Ph.D. Postlethwait is a professor of biology at the University of Oregon, Institute of Neuroscience. He studies the genetic regulation of animal development including development of the nervous system, the mechanisms of sex determination, the origin of novel morphologies in evolution and the evolution of the vertebrate genome.
Can you summarize the embryonic developments of the zebrafish, basically what you have been working on in your lab?
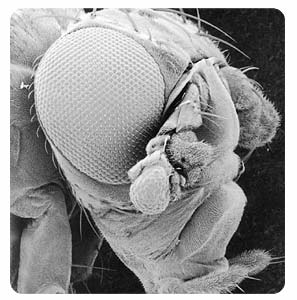
When I started my graduate work, one of the most fascinating things to me was a fly mutant that had legs growing out of its head. This was in the 1960s and this was before molecular genetics had really started. It was just really incredible to think of a fly who had legs where it should have an antenna. It’s growing a complete normal leg out of its head-two of them. And that was just fascinating to me how the developmental signals could get rearranged.
I began to investigate those things, many years ago, and tried to figure out how this mutation in a fly could change the fly’s antennae into a leg. What we found was that there were similar coordinates that told where the position was in an antenna, in a leg, and there were the same general properties that told anterior from posterior, back and front and proximal and distal in the appendage. And cells then decided that they were going to be leg cells or antenna cells. The appendage was sometimes a mosaic of those two types of cells. So clearly there was a genetic switch. And this mutation-it’s called antennapedia-had made a change in the gene that regulates a whole group of genes that cause a cell to be a leg cell or an antenna cell.
Subsequently, when molecular genetics really started going, other laboratories identified the molecular genetic nature of this gene — this antennapedia gene. It turns out that the gene encodes a protein that has a series of about sixty amino acids, called the “homeo domain,” which binds to DNA, and which controls the expression of other genes.
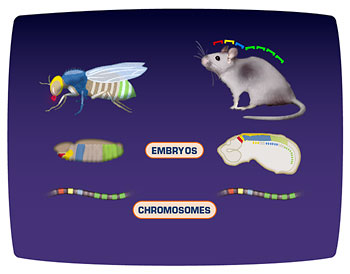
There are a number of genes in the fly that have this conserved group block of amino acids. These are in the Hox cluster, and the amazing thing is that humans have 59 of those 60 amino acids that are found in the fruit fly. In other words, this gene is very, very similar between a fly and a human. And this gene acts in humans and in flies to tell position in the body-to tell anterior/posterior position. This series of genes-this Hox cluster-controls how the body gets formed, anterior to posterior.
The genes are arranged on the chromosome in the same way that they’re expressed in the body. The genes not only control anterior to posterior but also in the limbs, they control proximal to distal. So you’ve got these 13 genes in humans that control these two dimensions.
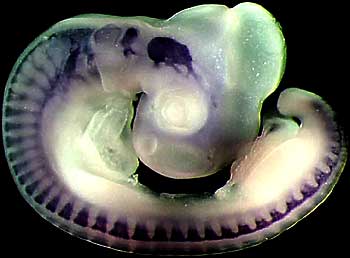
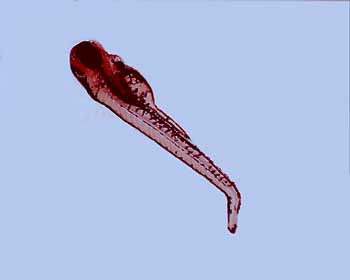
Eventually I stopped working on flies and started working on zebrafish. Other researchers had shown that these genes in mammals and in fish helped control brain development-the development of our hindbrain, especially, and our spinal cord. We were interested in the role of what sets up the signaling that tells the hindbrain whether it should be the front part of the hindbrain or the middle part of the hindbrain or the rear part of the hindbrain. The hindbrain is divided into segments, and these genes, these Hox genes are expressed in different segments, going back down the brain, and they regulate that.
We cloned a number of the Hox genes from zebrafish. In fact, we figured out the whole complex. To go back a bit: the fly has a series of Hox genes-about eight of them in a cluster. Ancient chordates-forerunners of fish, of frogs, of chickens, of humans-had a single Hox cluster with probably thirteen genes in them. In the vertebrate lineage, those duplicated twice to give one group of thirteen, two groups of thirteen, four groups of thirteen, and those are then four Hox clusters. Those Hox clusters are called cluster A, B, C and D. That’s true in human, mouse, chicken.
What we found in zebrafish is seven Hox clusters instead of four. So the zebrafish has two Hox A clusters, two Hox B clusters, two Hox C clusters, and at least one-maybe two-Hox D clusters. What had happened in the fish lineage is that these Hox clusters had duplicated again. Think about a lineage coming from an ancient organism to human, an ancient organism to fish. There were four Hox clusters back in this organism and then it duplicated to eight Hox clusters in the lineage giving rise to fish.
As we investigated we found that fish had probably a whole genome duplication. That means that about two hundred million years ago, the organism in the fish lineage, had twice as many genes as in the human lineage. Many of those genes have disappeared. The amazing thing is that the development is still very, very similar. Even though for many human genes, zebrafish has two copies, what we’ve discovered is that the zebrafish genes share the functions of the single human gene.
You also get very similar brain shapes and brain functions in zebrafish and humans, in the developmental mechanisms. The Hox genes work in very similar ways-even though there are two copies of many of the genes in zebra fish-as they do in humans.
Could you summarize what the benefits are of the current day tools that are available to researchers that study genetics today?
There are a number of methodologies and tools that are available today to genetic researchers and genomic researchers that were really unimaginable a number of years ago. One of those is called “in situ hybridization.” One thing that I would’ve loved to have known back when I was initially studying the antennapedia mutant in fruit flies is where certain genes are expressed in the body. In situ hybridization allows you to investigate where different genes are being used in the body.
All cells in the body have the same genes, but different cells in the body use different genes. So it’s a key question to figure out which cells are using which genes. With in-situ hybridization, you can make cells that are using a particular gene show up blue. In a zebrafish embryo, all of the cells in the embryo are visible and nearly transparent except the cells that are using a particular gene that you want to investigate-those show up as blue. This is a really fantastic technique.
Another technique is “Rapid DNA Sequencing.” This has allowed researchers to sequence the entire human genome and so all thirty-three-billion base pairs are now-nearly all of them-known. What this allows us to do then, is to very, very quickly move from a mutation or from a short piece of a gene to knowing the complete gene structure. And that’s really an amazing ability. That also allows us to go from a zebrafish genome to the human genome because we can compare-now as we’re learning the sequence of the zebrafish genome-that to the human genome and see how very, very similar they are. This comparing of sequences allows you to learn where on the chromosomes different genes are. Genes encode proteins, so you can identify the protein-encoding parts of the genes.
But also, importantly, if you compare the genome of zebrafish to the genome of human for a particular gene, and you look outside the coding regions-outside the regions that actually code for protein-you can find regions of a hundred or two hundred or more base pairs that have been conserved for 450 million years since the divergence of the human and the zebrafish lineage. Those regions that have been conserved have been kept similar by natural selection. What that means, then is that that identifies a region that is functionally important. That is, if you change the base sequence of the DNA in that particular region, you alter the function of the gene. And so those have been maintained.
So, along with Mark Featherstone, our colleague in Montreal, we compared a particular Hox gene sequence — the coding and the non-coding part –from zebrafish and mouse. What we found was that there was a particular part that was conserved in zebrafish and mouse. Then Mark took the zebrafish part and put it into a mouse and saw that the zebrafish part was expressed in the mouse exactly like is in the zebrafish. So then we were able to see which parts of the DNA were the same in the zebra fish and the mouse. Mark then made mutations that altered that and found that that altered the way that the gene was expressed. What that tells us is that we can use DNA sequencing to identify parts of genes that are conserved. Those are then candidates for regulatory regions and that’s going to make it easier for us to identify regulatory regions.
Can you briefly explain the process of cell differentiation?
Cell differentiation involves, as the word implies, cells becoming different. So cells become different from each other in space, and cells become different from each other in time. As time goes on, an individual cell will become different from what it had been before. And as cells divide, liver cells become different from eye cells, become different from pancreas cells, because they’re expressing different genes.
A key question is how cell differentiation occurs. How does a cell actually become different from what it was before — different from neighboring cells. This involves changes in gene expression. How do cells control gene expression?
Cell differentiation occurs because cells talk to each other. When cells find themselves in different environments, interacting with different cells, the cell signals between each other will be different, and then that causes changes in regulatory machinery inside the cells, which then ultimately causes different batteries of genes to be used in different cells. When different genes are used, different proteins are made in different cells, the proteins cause the cells to have different shapes and different functions.
Are all of an embryo’s cells always totipotent? If not, what is the process that happens to specialize the cell?
As development proceeds, the capacity of each individual cell decreases. That means the types of cells that it can differentiate into, decreases as time goes on. Consider a cell that has two progeny cells. These progeny cells have differentiated into two different functions. So one has one shape and one function, has one particular group of proteins. Another has a different shape, a different function and another group of proteins.
[J.P. points to top single cell in diagram illustrating cell division]
This cell is multipotent because when it divides it can give rise to both of these cell types. In contrast, these cells are not multipotent, and each cell here could give rise to only a cell just like itself. So how does this process occur? In the process of cell division, one cell can find itself in a different environment from the other. It can then express different genes and can send a signal to an adjacent cell. The adjacent cell receives the signal — there can be a receptor on the cell membrane than can receive the signal — and that then can be changed by a hierarchical pathway — a cascade of signaling to the nucleus — which then causes that cell to make a different set of proteins, which then causes a change in the cell shape and its function.
Can you talk about the function of Hox genes?
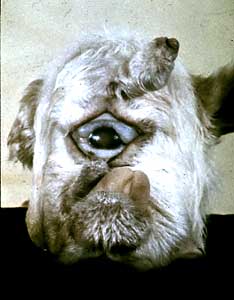
Hox genes encode proteins that regulate the expression of other genes. Hox genes control anterior/posterior patterning in flies and in people and in fish. So what controls the Hox genes? How do the Hox genes know to be expressed here, or here, or here? In fish and in people that’s controlled by a substance called retinoic acid. Retinoic acid is produced at high concentrations, about here in the body [gestures to back of neck/shoulder] and then it diffuses forward [moves hand upward alongside of neck to back of head and then forward to forehead] so the amount of concentration is lower towards the front end than it is back here [gestures back of neck/shoulder again]. And that tells which Hox gene is going to be used, which then tells which part of the brain, and which part of the face is going to be formed.
If you have a fetus that’s developing in uniformly high concentrations of retinoic acid you’d have then high concentrations not only back here where they belong, but also up here. What you get then is a baby whose face-and whose brain-doesn’t develop properly because it’s making posterior parts too far anteriorly.
The situation is set up when people take medicines that have high amounts of retinoic acid in them. For example, the common medicine Accutane is an effective controller of acne-because retinoic acid controls skin development as well as brain development. Accutane has retinoic acid activity and if a woman takes this medicine when she is pregnant, then the embryo is bathed in this high concentration of retinoic acid that controls Hox genes-in the wrong place. The baby’s face and brain don’t develop properly and the baby is born with abnormalities.
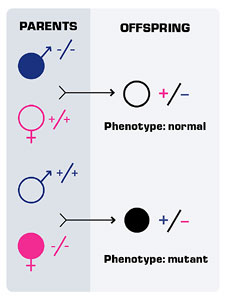
Please explain what caused malformation in these zebrafish embryos in regards to the Hox gene?
We know that retinoic acid controls Hox genes. You can take a zebrafish embryo and grow it in media that contains retinoic acid. It causes two things that you can see. It causes the Hox genes to be expressed more anteriorly than normally they would and also it causes the parts of the face of the fish to develop abnormally.
Can you talk about cell signaling?
Cell signaling involves one cell sending a molecular message to another cell that causes the flipping of genetic switches in the recipient cell and that alters that cell’s activities.
Can you describe the concept of the “genetic toolkit”?
Because different organisms look so different-a fly looks so different from a human-we used to think that their developmental regulatory mechanisms must be very, very different. However, we have discovered that the genetic took-kit used by a fly and by a human and most other animals is very, very similar. So a genetic toolkit is the basic groups of genes that control how developmental processes occur.
For example, Hedgehog is part of the toolkit, Hox genes are part of the toolkit and they’re used in flies and they’re used in humans in very, very similar ways.
It’s utterly amazing to me that similar genes can control development in such a wide variety of creatures. For example-look at the face of a fly. And then look at your own face. To think the same processes are controlling the way a fly’s eye develops, the way a fly’s antennae develop and the way your limbs develop, the way your eyes develop, the way your brain develops-it’s utterly mind-boggling.
It’s truly amazing how similar the action of genes are from very, very different species. For example, in fruit flies there’s a mutation called “eyeless,” so the flies with this mutation don’t make an eye. You can isolate the same gene from humans. In humans when that gene is mutated, it causes a person not to make an iris to the eye. And if you take that gene from a human and put it into a fruit fly cell and cause that cell to develop into an eye, it makes an eye. So that the human gene can replace the function of the broken gene in the fruit fly.
Does it make it a human eye? Or a fly eye?
It turns out it makes a fruit fly eye. The human gene expressed in a fruit fly cell causes a fruit fly eye to be formed. And that’s because what causes the eye to be formed is the downstream targets. So that the eyeless gene is a regulatory gene-it turns ‘on’ different genes that make the actual eye. Those genes that make the actual eye are still fruit fly genes, not human genes. What the human gene does is it turns ‘on’ the genes those fruit fly genes.
Can you talk about why certain species have been selected to be used as model organisms in research?
There are a number of organisms that researchers focus on in order to understand how organisms develop. These are called model organisms because many researchers all over the world use these organisms. Examples are fruit fly, a certain nematode worm called C. elegans, the zebra fish, mouse, frog. Those are all models for human development and they’re used because they develop rapidly, they have genomes that are easily manipulated, their embryos are easily manipulated by a researcher-you can take them outside-they don’t develop inside the mother except for the mouse, and they’re easy to study for a researcher in the laboratory.
What are some of the big questions that you have regarding your research and just genetics research in general?
A big question, then, is how-if we have the same basic genetic toolkit for flies and fish and humans-can the basic genetic mechanisms be used to make animals that look so very different? So initially biologists focused on the differences between organisms. Now we’re amazed at the similarities-at the genetic level-among organisms. But still we have to now go back and understand how those genetic similarities become manifest to make the differences among organisms.
We are on the threshold of being able to understand how evolution has produced organisms that look as different as flies and people. What we need to do is to compare the activities of genes-not only between organisms that are as differently-related as humans and flies, but also of organisms that are more closely-related that also differ in shapes. For example, different fish that can interbreed, but yet have very, very different body forms-how the genes control those different body forms.