





Rediscovering Biology: Molecular to Global Perspectives
Neurobiology Expert Interview Transcript: Fred Gage, Ph.D.
 Professor of Genetics
Professor of Genetics
Fred Gage, Ph.D., is a professor in the Laboratory of Genetics at the Salk Institute in San Diego. His research is focused on the generation of new neurons during adulthood, an idea that runs counter to neuroscience dogma. Gage hopes that the work will one day lead to therapeutic uses for people with stroke, paralysis, or Alzheimer’s disease.
Interview Transcript
Fred Gage, Ph.D., is a professor in the Laboratory of Genetics at the Salk Institute in San Diego. His research is focused on the generation of new neurons during adulthood, an idea that runs counter to neuroscience dogma. Gage hopes that the work will one day lead to therapeutic uses for people with stroke, paralysis, or Alzheimer’s disease.
Explain neurogenesis.
So what is neurogenesis? Neurogenesis is really a process rather than a single event. It involves the cell division of what we call an uncommitted cell. It stops dividing and then it begins to make choices as to whether or not it’s going to become one of any number of types of cells within the brain lineage or neural lineage, and it can become a glial cell, of which there are two types, astroglia, and oligodendrocytes, or it can become a neuron, or it can die. And if it chooses a neural phenotype, then there are literally thousands of different types of neurons that exist in the brain.
So in the process of neurogenesis we have cell proliferation, cell determination, cell fate choice, as we call it, and then once the cell makes a decision to become one type of cell or another, then the process of maturation and differentiation begins, and this goes through to the point that the cell is integrated within the circuit and is functional. So from our perspective, neurogenesis incorporates all of these processes from birth to full maturation.
Define a stem cell in general and the stem cells that you work with.
There’s one good definition for stem cells. A stem cell is a cell that self-renews. That means it can give rise to itself over and over again and can, upon division, give rise to a daughter cell, so one of the cells that the cell divides into is itself-self-renewal-and the other is a daughter cell, which then can differentiate into some sort of other cell, and that type of mature cell is different from the stem cell.
Now, the underlying assumption is that it can give rise to a variety of cells, but that’s not really implicit in the definition. It just means that the cells are self-renewing and that one of the daughters can give rise to a variety of other cell types.
But there are many different types of stem cells and they’re characterized differently. The most primitive stem cells are what are called “pluripotent” stem cells, and pluripotent stem cells are cells that can give rise to all the different lineages of the body, in fact, and these cells are also called ES cells or embryonic stem cells, around which there’s significant controversy right now in terms of the human use of these cells.
There is only one cell more primitive than an ES cell, which is a fertilized egg. That cell is what we call “totipotent,” because that cell, when implanted in the womb of a woman, can give rise to the entire body, a baby, whereas the embryonic stem cell cannot. It is missing certain support factors that allow it to form organs. So while embryonic stem cells can give rise to all the cells of the body, they can only give rise to them as individual cells and not as organs. So the problem of organogenesis is something that requires a separate factor from the embryonic stem cells, which we really don’t understand yet.
So a totipotent cell is the fertilized egg and pluripotent cells are the embryonic stem cell. As development progresses from the embryonic cell, which is derived from the blastocyst of an embryo, these cells are isolated from the blastocyst and are put in a culture where they can divide. And those cells then can give rise, as I said, to all the different cells of the body and there are three major lineages that are important-the ectoderm, the mesoderm, and the endoderm-and these three divisions make up all the cells of the body, and ES cells can give rise to all of them. If the embryonic stem cell stays within the embryo and it implants, it will evolve, or the blastocyst will become an embryo, and the embryo will develop and at an early stage of embryonic development is something called “organogenesis” where presumably these embryonic stem cells migrate to regions where they seed the beginnings of new organs. And this is believed to be the period of time that the totipotent cell becomes a multipotent cell. And this is a cell which we believe to be lineage-restricted, so at this early stage, when the cells migrate to form new organs, we can have stem cells of the liver, we can have stem cells of the blood system, stem cells of the brain and these cells are the origins of these structures.
And these cells we are finding now surprisingly persist throughout life in these organs — albeit at very, very low frequency-but now we believe that every organ in the body has persistent stem cells that are for the most part quiescent, but with any luck we might be able to learn enough to be able to reactivate them under certain circumstances.
So the lineage then is totipotent (the egg), pluripotent (embryonic stem cells), and multipotent (lineage-restricted cells). And within that third domain, there are neural stem cells, and these are cells that reside within the ventricular zone, or the area near the closure of the neural tube where they give rise to neural progenitor cells. So it is the neural stem cell that continues to divide and gives rise to the neurogenic event that we’re examining, and those cells then slowly divide, and with age there are fewer and fewer of them, but again the surprise is that they persist throughout life.
Explain the importance of the hippocampus.
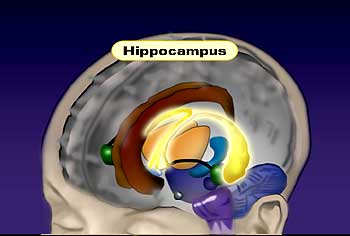
The hippocampus is a structure in mammalian brains that resides within the temporal lobe. It’s surrounded by the temporal cortex; it’s embedded in there. The hippocampus is actually as you know a seahorse, and the shape of the hippocampus is analogous to the seahorse. It has this same sort of U-shaped or curved shape to it. And it has a very distinct anatomical organization, a very distinct pattern.
It’s called the archaecortex because in very low organisms in evolution, this was what existed as the cortex. For example, in lizards this was the major cortical area, and then as the neocortex or new cortex evolved, the archaecortex, which is the hippocampus and its surrounding structures, became more of a subcortical structure in many ways. So the hippocampus and its surrounding cortical structure in the temporal lobe is considered the archaecortex or the ancient cortex, which was the first evolved cortical structure and then as the neocortex evolved, the hippocampus and its structures actually are now not thought of as cortical structures but as subcortical structures.
Functionally, the hippocampus is thought of as an area of the brain involved in learning and memory initially, and some of the key studies that tried to unravel this function emerged from Brenda Milner, a colleague who works at Montreal Neurological Institute. Patients with epilepsy, which strikes the hippocampus in some cases, were treated by surgery where the hippocampus was the focus of epilepsy, and surgeons would remove the hippocampus. And if you unilaterally take the hippocampus out, there’s very little deficit in individuals, so it appeared to be a safe operation to do this.
In a few cases, in a very famous case of a patient called HM, both hippocampi were removed, and some of the surrounding tissues and that patient showed a very distinctive memory loss of recent memory. So all old memories he remembered. He remembered everything that happened to him as he was developing but could not consolidate or form new memories after the removal of both hippocampi.
So the current thinking about the function of the hippocampus is built on that original idea that hippocampus is involved in processing new information and it really can be thought of as aggregating information from all the sensory and motor and emotional domains of the brain into one structure.
So, for example, in this setting here we have visual input, we have auditory input, there’s some emotional impact if what I say has some significance for you in some way. You’ll have some tag of emotion on it. There are motor events that have to be taken into consideration.
All of these things-even the temperature of the room. All these sort of things are weighing into what will be a memory for you of this event subsequently. So all of these different sensory, motor, and emotional domains are coming into the cortex. That information is then passed through the hippocampus where we believe that information is aggregated in some way and then it is stored back out in the cortex as an aggregate.
So long-term memories are difficult to retrieve as individual sensory events. It would be difficult for you to remember this meeting just as my face and not anything associated with it, and obviously memories-the different sensory motor components are weighted differently, so hopefully the content of what I’m saying, the verbal content, is weighted most heavily, and that will be remembered most, but in other circumstances there’s very little verbal input, and most of the memory is composed of visual input, so it’s really a weighting of these different events into the memory package but they all are integrated.
So the important point is the hippocampus is not a storage site for information but rather a processor of recent memories where they can be then stored into the cortex where they are stored presumably in a distributed way.
What does the declaration of the 1990s as the decade of the brain mean to you?
The decade of the brain, 1990 through 2000, I think it indicated a period of time where neuroscience as a field had matured to the point where the information was gathering at a very, very rapid pace, in part because the tools were available to acquire information and relate that information to other existing disciplines as well. And I think two major technical advances occurred as a thread through many of the other advances.
The first is the molecular mechanisms underlying brain function, so the cell and molecular study of the brain became doable as a study. The tools were available, and no longer was the brain really considered as much of a black box. And the revolution there comes in part from everyone in biology because of the ability to clone and sequence genes and to get access to molecular information in many different other areas. So neuroscience was taking advantage of that for the first time, and this continues to have an enormous impact on neurobiology. And so we’re really not using any other tools than are used in other areas like cancer biology, but by applying them to the brain we’ve really brought the brain into modern era.
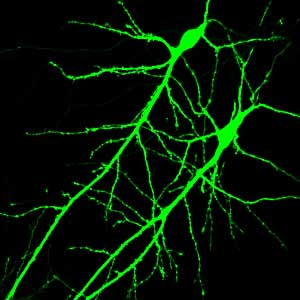
The other major development was in imaging, so by imaging I mean this at a variety of levels: there is microscopy allowing one to look at very, very small parcels of the brain using very specific markers to indicate differences between cells or differences between compartments within cells. And examples of some of these kinds of tools would be confocal microscopy, which has evolved into multi-photon confocal microscopy. Other methods, of course, are related to larger structures, and in this way, one can utilize digital imaging and begin to see cells that are being examined in real-time so watching them on a temporal basis.
So the first thing I said was that you can look at small objects and label them using special fluorescent tags and microscopic techniques and characterize them very carefully and very accurately. The second bit is being able to monitor these brain cells in a temporal domain, so we don’t see it as a static structure anymore but a dynamic structure.
And these are all tools used a lot for molecular questions, but imaging also is reflected in magnetic resonance imaging and positron emission tomography and their prodigy, and the other kinds of techniques that have been developed that allow clinical investigators and human investigators to image the brains of actively behaving or thinking or performing humans, and this has caused an additional revelation and revolution in our way of thinking and way of experimenting. So I think between the molecular tools and the imaging tools, the ’90s have really been a real data accumulation period of learning.
What did people used to think about the growth of neurons prior to the developments in your work and what you found out about neurogenesis?
I think it’s fair to say that certainly in the first half of the last century and well into the second half, there was a belief that once the process of development has ended-postnatal-presumably there was no substantial amount of new neurons that were born into the brain. And yet there was significant study of neurogenesis during development, so there’s a rich literature on embryonic and neural development in the embryo.
But in fact it was in the early sixties that we had the first indication that there might be some cell division continuing in the central nervous system, and these studies were using methodologies that used radioactively labeled thymidine as a marker, which was incorporated into the genome, the DNA, of a dividing cell and would act as a persistent marker throughout life. So you could give a single injection of this non-toxic compound and then look at various times after that injection to see whether or not there were dividing cells, and then, secondly, did they persist, where are they, and then more importantly what did they become. Were they just dividing and dying, or did they continue to divide?
There’s a Joe Altman who used these studies with thymidine to demonstrate that there are dividing cells in the brain, and this period of time was actually very exciting. People were excited by the idea, but in truth it’s likely that the reason it did not catch on was because the tools were not definitive, so it could be that there were so many other explanations for what might be going on and the ability to prove that the cells had actually turned into neurons was not as good as it could be or would be in the future.
In addition, I think there were no studies of quantitative biology at that time, so you didn’t really have a sense of how much this occurred, and I think some people said, “well, if it occurs maybe it’s at such a low rate that who really cares. It’s just can’t be that important.”
In addition, there was this general idea that the analogy for a brain’s function is that of a computer: that it’s a fixed process and memories are stored in these circuits, and how could you possibly add new cells into the circuit and not disrupt them in some ways? So this was an untenable argument that there are new cells being born and integrated into the circuitry.
In the middle eighties, Fernando Natibom and Steve Goldman from Rockefeller demonstrated using this thymidine technology that songbirds could generate new neurons, and that it was seasonal and it occurred during the acquisition of new songs each year, and that was very exciting for people. But I think even at that point it was considered to be an avian phenomenon, and while it was interesting for birds it probably had no impact on what was going on in mammals.
Again, all that was slightly problematic due to the methodologies and the ability to confirm that it actually turned into neurons, and it really wasn’t until the early nineties that technology had advanced far enough so that one could use, for example, confocal microscopy along with antibodies which specifically stained for elements of neurons that were convincing, and then you could multiply label the cells. In addition, another analog to thymidine came along, bromo-deoxyuridine, for which there was an antibody. This allowed you to look more specifically at the cell itself. The thymidine was a radioactive marker, so the radioactive particles that would come from the cell that was carrying the thymidine were covered with emulsion, so the visualization of the particles was always slightly distant from the cell itself, whereas the antibody is very specific for the nucleus and could be stained and amplified using amplification techniques.
How were the findings in songbirds ultimately replicated in humans?
So the songbird work was not in the hippocampal structure. It was in a song area of the brain, and, as I said, that whole story evolved separately as an idea that there was some self-renewing that was going on in this one structure involved in the acquisition of new songs each year. But the relationship of the replication seen in the songbird to that seen in the hippocampus of mammals has never really been made because that’s not the way in which it happens in mammals. There’s not an annual or seasonal turnover.
So actually after ’83-’84 when that came out there was actually quite a lull again like we saw after Joe Altman’s period in the sixties where there was a burst of enthusiasm and then it sort of faded out of focus. And partly I say that was because of the technology. Even in the songbird work, the questions about whether or not there were authentic new neurons and how many there were and whether or not they were actually structurally wired up appropriately, and what the mechanisms were, these were all still being resolved.
But, again, in the early nineties some new technology evolved that allowed us to confirm in the mammalian brain and specifically in rodents-mouse and rat-that in the hippocampus and in the sub-ventricular zone, two areas of the adult brain where cell proliferation clearly continues, there was evidence that cells could take up a marker called bromo-deoxyuridine, which is an analog of uridine, into the DNA and now you could use antibodies to stain the nucleus and antibodies to stain the cytoplasm or the cytoarchitecture of the cells to confirm that the cell that had been born at the time that the bromo-deoxyuridine was injected had, over time, developed into a mature cell.
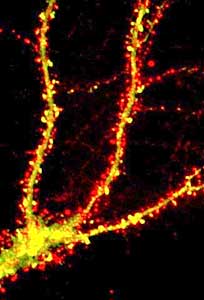
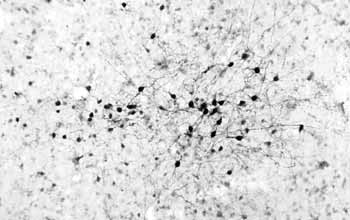
In addition to these confirmations using bromo-deoxyuridine, confocal microscopy was a really important element to this whole process because regular microscopy looks at thick sections, and there was some concern that the bromo-deoxyuridine staining may not be inside the cell but may be on top of the cell and the surface of the cell or two cells adjacent to each other. And what confocal microscopy allowed one to do was to optically dissect the tissue with a laser and take digital pictures all the way through the cell and then reconstruct the cell in three dimensions to prove that the cell that had been labeled with bromo-deoxyuridine had over time differentiated into a fully mature cell.
So the confocal microscopy really nailed it down that neurogenesis was occurring and the significance of it was further amplified by what is called “stereological quantitation,” so these are methods that have been developed over the last ten or so years that can accurately count how many cells of a certain type exist within the brain. The brain has got so many cells and they’re so densely packed that it’s difficult to really get an accurate representation of a subpopulation, but with these good marking techniques and this quantitative methodology that could be superimposed on that, it became clear that there actually were a lot of cells that were being born in the adult brain throughout life.
That raised interest because it wasn’t just a passing phenomenon and it was also shown that they persisted throughout life, so they weren’t just born and died off. They actually persisted.
Armed with that, more people started looking at this in a real way. Our work in rodents developed the technology, and there were several groups that were looking at primates-at monkeys, nonhuman primates-including Bruce McEwen’s group, Fuchs in Germany, and Gould who was a student of McEwen, as well as at Yale, Pascal Requicha. And they were all using similar techniques that had been developed in the rodent to see whether or not it occurred in the primate.
A postdoctoral fellow in my lab at the time, Peter Erickson who was from Gothenburg, Sweden, was part of a group of scientists in my lab thinking about ways in which we could figure out whether or not we could determine if neurogenesis occurred in humans.
And several of the folks in my lab were clinically trained, and a couple of them were neuropathologists, and we reasoned that, based on some thinking about it that the experiment in humans had already been done, and that is because bromo-deoxyuridine had been given to patients that had cancer. And then what would happen is that at various times after the bromo-deoxyuridine injection, a biopsy would be taken from the presumed cancerous organ, and then you would look at that tissue, the biopsy, to see how many of the cells were actually dividing as an index of how metastatic or how cancerous the tumor was. So it’s a diagnostic, not a therapy but a diagnostic.
But bromo-deoxyuridine is actually transported through the blood. It’s a very small molecule, so in addition to labeling the tumor, it would get to the brain and label the brain, because that’s how we give it to animals is through the blood.
So we figured that maybe we would be able to just look in the brains of pathological samples that already existed, so people that had cancer that had been tested with BrdU that had died. I called a bunch of my friends who were pathologists and got samples of sections of brain from patients that had died, and we actually saw BrdU labeling in the brains of some of these sections, but we could never do confocal microscopy, quantify it, or really do double labeling to confirm that they differentiated into neurons, but we were heartened by the idea that they were there in humans, at least they were dividing cells. But we realized we needed fresh tissue or fresher tissue.
So Peter went back to Gothenburg, and several other fellows went back and tried to get involved in ongoing clinical studies so that we could get fresh tissue. Peter got involved in a cancer study that allowed us to get the tissue from patients that died within a short time earlier.
And the nurses and the entire team there worked very diligently with the families and with the protocols and were able to get us very fresh tissue, and over a period of about two and a half years we got five or so patients that had good quality material, and we were able to then use the methodologies that we developed accurately in tissue in live animals, and proved then that the newly born cells in the adult human brain had differentiated into neurons.
Interestingly, the patients that we had in the study were all in their late fifties up to age 70, so we actually showed that neurogenesis, as indexed by these tools, persists throughout life in humans, and even in ill individuals one sees this kind of neurogenesis occurring.
Interestingly, that came out right about the time that the studies were coming out in primates, too, so there was this period between 1997 and 2000 where human and nonhuman primate studies were confirming that neurogenesis occurred.
How was the connection made so that you could return to the use of mice as an animal model to look at neurogenesis now in your lab?
Being able to investigate really how this occurs and what regulates it and how it might be useful is not doable in humans because of this heroic way in which one had to get this tissue, and we were not prepared really at these early stages to use a lot of primates for the basic studies.
But what was clear was that the process of cell division just within the hippocampal zone and the movement of the cells and the differentiation appeared to be analogous from mouse to human, so in all species that we looked at and others had looked at, the process of neurogenesis looked very similar, which gave us a lot of confidence that we could go back to experimental animals and learn more about the fundamentals of it.
Certainly, as new surprises arise you want to then periodically try and confirm this if you can in humans and primates as much as you can, but most of the fundamental work comes out of the work in mice and rats.
What are the steps of neurogenesis and what factors might have influences along the way?
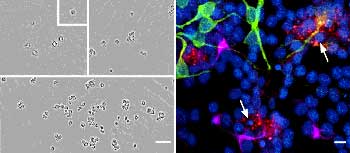
So now a lot of effort is going into trying to understand just exactly how neurogenesis functions mechanistically. Now one of the great advances in the field has been the ability to develop the techniques for isolating cells from the adult brain, for isolating neural stem cells and looking at them in culture. So we can take now from the adult hippocampus of all species in fact, from mouse up through human, we can isolate the cells in culture and we have found molecules, growth factors and cytokines, molecules that induce cell division. Mitogens are generally the category of these molecules, like fibroblast growth factor and epidermal growth factor are two distinct molecules that can maintain the division of these cells in culture. So we can grow up millions of these stem cells in culture, which affords us the opportunity now to examine cellular/molecular events that are involved in the process.
To that end, we and many colleagues have discovered not just that we can induce cell division, but understand more now about the differentiation process and differentiation really is only one of the choices of these cells, because we found that both in vivo-that means in the live tissue-as well as in culture, a large number of these cells that divide give rise to mature cells, but another large percentage actually die off. So dying off or cell death is likely a very important selective process in this whole mechanism of giving rise to new neurons to control the number of neurons that are coming into the circuitry.
We are beginning to understand some of the molecules that are responsible for inducing differentiation but one of the more interesting observations that we’ve made recently is to think about neurogenesis not as a cell autonomous event, that is, not that everything is programmed within the stem cell itself, but the stem cell is making its choices in the context of its environment. So we’ve seen both in the brain and in our culture dishes that you can co-culture or look at the environment the cell is in and recognize the fact that subpopulations of support cells are actually critical for making the decision of the cell to become one type of cell or another.
So, for example, glial cells, which are one of the lineage choices for a stem cell, in and of itself can tell a stem cell to become a neuron. So when the stem cell’s dividing in proximity to certain types of glial cells, they then become neurons. Interestingly when stem cells are growing in the region of another type of cell like another neuron, then it becomes a different type of glial cell, an oligodendrocyte, so we’re beginning now to think of this neurogenic niche as being critical for the choices.
So there are cellular niches, which are these cell types that interact with each other, and then there are the molecules that are on the surface of the cells as well as molecules that are secreted by these cells to affect this occurring, and that’s really where the field is right now, is finding the molecular events and the cellular context within which those events are occurring.
There are candidate genes for all the different processes-migration, differentiation-but I think the truth is it will not be any one molecule that defines this but a variety of molecules that can affect it, depending on the context in which they are being presented or the cells find themselves.
What are the niches where stem cells exist, and where can others migrate to, and what’s the functionality of the cells once they have migrated?
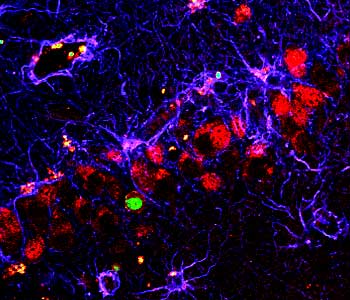
There aren’t niches that exist in certain areas of the brain like the hippocampus where neurogenesis can occur. But surprisingly there are dividing cells that exist throughout the brain and spinal cord and this is actually something that is a big surprise: that the spinal cord, the cortex-all these other structures of the brain-they have dividing cells in them, but the current view is that they don’t have the appropriate niche or the support structure to allow them to become neurons.
And there are two ways that you can interpret this actual difference between the stem cells in different regions. You can say that there’s something intrinsically different about the cell, or you could say that the environment is different in these two structures, and that’s a current debate — not really debate but area of investigation — how much of it is one vs. the other-nature vs. nurture. The answer is probably a little bit of both, but certainly it’s important to examine their clear differences in the structural environment in which the cells occur, and you can isolate stem cells from an area where no neurogenesis occurs and transplant them into an area where there is a niche for neurogenesis, and those stem cells can become neurons. So that suggests at least that these other areas where no neurogenesis occurs is not filled with cells that can’t become neurons. It’s just that the environment doesn’t allow that to occur.
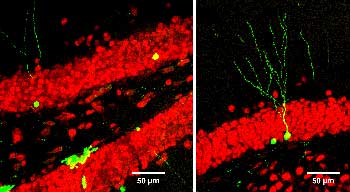
Now one of the fundamental questions that is sort of hidden behind this is, can these adult cells that divide and presumably differentiate, do they actually differentiate and become real functioning neurons in the circuit? And that’s been a question that has eluded us for many years, and just recently we’ve developed some techniques and published some result directly at that.
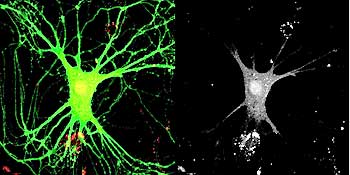
And what we did was to take advantage of a tool in molecular biology, which is a virus, a retrovirus, which has the capacity of infecting only dividing cells, and we mutated the virus so that it was not infectious, it was not proliferative, so once it infected a cell, it wouldn’t propagate its infection around to other cells but would stick within that cell.
And we engineered it in such a way that it expressed a bright green color normally, and our strategy here was that we wanted to label the newly born cells and track them throughout life and then be able to record from those cells with electrophysiological techniques, specifically patch clamping, so we could patch onto or electrically record from that newly born cell and ask whether or not it had developed into a mature cell.
So over a period of years, several individuals in the lab including importantly post-doctoral fellows Henry van Praag and Alshondo Schinder conducted a series of experiments that demonstrated that newly born cells in the adult can differentiate into cells that are indistinguishable from sister cells that had been in the brain for long periods of time. But importantly, the process of differentiation and maturation, which is after they had grown their processes and they actually have electrical properties that are mature, they continue to change over months and months. So our current view of it is that these new cells that are being added into the circuitry are really young cells that maintain the flexibility or adaptability of the hippocampus throughout life because these cells tend to have maturation ongoing all the time sort of seeded throughout the hippocampus, and this adds an element of immaturity to a mature hippocampus. So that’s our current view.
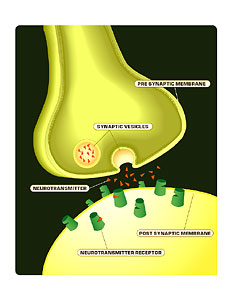
But importantly we have shown now definitively that the cells can mature into functional cells with electrophysiological properties, and they interconnect with existing neurons and they make signals and release transmitters.
What is the role of the enriched environment experiments with the mice?
One could think about neurogenesis as a static event. That is, that you have this added number of cells coming into the circuit and that affords an individual some function of which we don’t really know yet. We’ve now found that they are functional. What is the function that they have? One could theorize that they are continually added into the circuit. We had a surprise that occurred, ah, some five or six years ago where one of the fellows in my lab, Gerd Kemperman, was reading the old literature on environmental enrichment where they showed that there were changes in synapses, changes in transmitter release, changes in neurotransmitters, and he questioned whether or not the events of neurogenesis were modified by changes in environment, and really it was a wild shot. It was not something one could predict ahead of time, but the effect was quite robust. So if you take genetically identical, sex-matched to the same gender, same age adult mice and then split them into two environments, one which is standard laboratory conditions, impoverished as you might call it, vs. the littermates or the siblings which are placed in an enriched environment with toys and much more space and things, and they interact for merely 45 days and then we look at their brains.
We found that there were as many as 15% more neurons in this area of the hippocampus in the enriched group as compared to the control, and this increase in numbers of neurons was correlated with an improvement and ability of the animal to subsequently learn new tasks, so you had an increase in neurogenesis and an increase in functional outcome.
When we looked carefully at it in the enrichment, what was happening was that there wasn’t an increase in proliferation of cells, but there were fewer cells dying off, and by having just fewer cells dying off we increased the number of cells that survived and integrated into the circuitry.
In subsequent experiments, we tried to tease out what elements of the environment might be important and one theory could be that just because the hippocampus is involved in learning and memory, maybe that was what’s going on. So we did a series of additional experiments where we separated out training the animal, but bringing them back to their home cage instead of having the enriched environment.
And as a control we just put a running wheel in the animal’s cage to control for just generalized activity that they would have had in the enriched environment but not in their standard cage environment.
And a second surprise came up, which was that just having access to a running wheel-so voluntary running in a running wheel-doubled the number of cells that were dividing in the hippocampus in a very short period of time. And this has subsequently been replicated by many individuals, and it is almost an assay that we use for increasing enrichment, so voluntary exercising would be a big boost, and it also supports survival and it has a very dramatic effect on learning and memory. So just voluntary activity has a very dramatic effect on neurogenesis. Part of our effort now is to try to unravel these correlations to see to what extent they’re actually mechanistically linked and what it is about environmental stimulation that causes this increase in neurogenesis, and whether that increase in neurogenesis actually causally related to the improvements in performance that we see in all animals that go through these regimes.
Can you talk about your ideas of the ability of the brain to recover function, to regenerate itself, and what sort of clinical applications there are for harnessing this power of the brain?
I think that there are a couple of implications of the direction that we’re going and thinking now about the brain. One relates to just normal brain function, and that is that the brain is a much more dynamic structure than we ever thought before, and that our behavior and our interactions in the environment can affect the structural plasticity, the structural organization of the brain itself and the structural organization in the brain actually controls behavior.
So as a neurobiologist I believe that the brain is the controlling organ for our behavior, but now we’re finding out that behavior can change the structure of the brain. So what this means is that who we are is affected by what we do.
We’ve always known that in some sense, but now we’re finding that to the extent that the hippocampus and its related structures in the organism are involved in the acquisition of new memories and the consolidation of new memories, and that is such an important part of who we are and how we interact in the environment. That is affected by what we do, so who we are, in many ways, is very much affected by what we do, and it’s a bit of a “do-loop,” if you will, where the brain controls behavior, but behavior then can feed back and regulate how the brain then will come and respond to that next environment.
And what it means is that we have actually a lot more control over who we are as individuals based on the choices that we make and the kinds of behaviors that we chose to do in our lives. So that has a sort of philosophical effect on our approach to thinking about the brain’s function and its relationship to behavior.
But in addition, one has to think about this also in terms of disease and brain injury, and from our perspective, the idea is that there are cells that exist, a residual pocket of cells that exist, that can divide and in some areas they can give rise to neurons.
Now one view would be that these regions like the subventricular zone and the hippocampus, where cells are dividing and giving rise to neurons, the cells might be able to migrate to an area of the brain where damage has occurred where they could then replace cells. If we could learn enough about molecules that are involved in migration and proliferation, and then maybe interface that with what we know about how activity influences the maturation and survival of the cells, we might be able to put up together certain kinds of strategies that would take advantage of this residual plasticity.
There is, as I said, very little spontaneous brain repair; the brain doesn’t repair itself very readily. But as we learn more about the mechanisms that do this, I think it could-and we’re seeing it now already in experimental animals where certain growth factors can be injected into the brain and induce the cells to divide and migrate to areas and repair them.
How soon this will actually get translated into clinical therapy is not really clear, because many of these molecules have many different effects, and we have to look for very simple compounds that have known effects. But it is quite clear that this is a direction that this field is moving in, and I’m very optimistic that in the near future there will be strategies for inducing what we call self-repair within the central nervous system by taking advantage of these endogenous cells that exist.
What questions are you left with?
We still don’t know what the function of these newly born cells is. We know that they’re functional, but what is their function? What role do they play in normal function? And we are working through a variety of techniques to try to figure out how that occurs. We’re very much interested in this concept of a niche, and what exactly are the cellular and molecular events that control the fates of these cells into the different lineages. How we can address that in some meaningful way?
And we look at that in culture dishes where we isolate the cells and try to understand how they make their choices, how they simply become one type of cell vs. another. That is, I think, going to take us certainly for the next number of years into the future. I think one of the main stories and issues that has arisen in my own mind as a result of the work is that the analogy of the brain as a computer, as a fixed organ in the adult brain, is probably not adequate or not a complete enough picture of what we’re learning about how the brain functions. The brain is as clearly an organ that is changing and modifying itself throughout life, and importantly organisms, humans, have the opportunity to participate in the change of that brain and the change in that function of the brain throughout life.
And since the brain really is the organ that controls behavior, this interplay between the structural plasticity of the brain and the way we interact with our environment is a different way of looking at the brain than the traditional view of the brain as a fixed circuit.