


Rediscovering Biology: Molecular to Global Perspectives
Online Textbook and Video
This online textbook chapter supports and extends the content of the Genetically Modified Organisms video. The chapter provides details of the procedures used in genetic modification as well as an in-depth analysis of the ethical issues surrounding the issue.
1. Introduction
“And God said…let them have dominion over the fish of the sea, and over the fowl of the air, and over the cattle, and over all the earth, and over every creeping thing that creepeth upon the earth.”
– Genesis 1:26 The Holy Bible
“The Earth does not belong to us. We belong to the Earth.”
– Chief Seattle
Introduction
Humans purposefully manipulate the evolution of other organisms. For thousands of years farmers have used selective breeding to improve their livestock and crops. As a result, we have cows that produce more milk, hens that lay more eggs, sheep with better wool, and disease-resistant plants with higher productivity. Another striking example of humans alting other organisms is the great diversity of dog breeds, from the toy poodle to the Great Dane.

The ancestor of modern corn had tiny kernels, each protected by a tough husk. Domestication of maize, which began thousands of years ago, selected for large sheathed cobs containing large kernels without husks.
Early maize cobs from the Tehuacan Valley. Fig 122 in the Prehistory of the Tehuacan Valley, V.1 by Richard S. MacNeish © Robert S. Peabody Museum of Archaeology, Phillips Academy, Andover, Massachusetts. All Rights Reserved.
Although humans have been manipulating organisms for millennia, genetic engineering simplifies and targets manipulations in an unprecedented way. Transgenic plants and animals are generated with characteristics that cannot be obtained using traditional breeding. Unlike organisms generated by selective breeding, transgenic organisms">transgenic organisms (also known as “recombinant organisms”) by definition contain genes from other species. Genetic engineering techniques are used to generaterecombinant DNA">recombinant DNA, which contain sequences from different organisms. This DNA then becomes incorporated into a host so that it can be passed to subsequent generations. For example, Bt">Bt corn expresses a gene for an insecticidal toxin that was “donated” by the bacterium Bacillus thuringiensis.
The use of recombinant organisms has become commonplace. For example, bacteria produce human insulin and hepatitis vaccines, and some crop plants are cultivated to be resistant to certain herbicides and insects. There are also transgenic livestock that produce human proteins, such as antithrombin III. The economic value of such products drives research. How did it start? Where is it going? What are the challenges and the risks? In this unit we will explore these questions as they relate to modifying bacteria, plants, and animals.
2. Genetic Modification of Bacteria
Bacteria – the first organisms to be genetically engineered – are used for replicating and altering genes that are subsequently introduced into plants or animals. Bacterial systems lend themselves to genetic manipulation in part because of their rapid reproduction rates. It is easy to produce a genetically identical population – a clone of bacteria – all containing the gene of interest in a short period. The cells can then be lysed and DNA can be isolated in short order. Bacteria are routinely used to produce non-bacterial proteins. An example is the production of purified proteins for vaccine use. Such proteins can be safer and as effective as vaccines that contain killed or attenuated (weakened) pathogens. Genetic engineering can also produce extensive changes in the bacterium’s metabolism. For example, bacteria can be provided with several genes, encoding enzymes that allow the production of fuel alcohol from wood.
Researchers have taken advantage of nature to modify bacteria. Plasmids are small, circular, self-replicating, extrachromosomal pieces of DNA that occur naturally. A plasmid can encode a protein that offers its host a selective advantage. For example, a plasmid that encodes an antibiotic allows its host bacterium to thwart competing microbes. Alternately, a bacterium might possess a plasmid that encodes antibiotic resistance. Plasmids are readily isolated from bacterial cells and can be altered in vitro by inserting or deleting specific sequences of DNA. Because they can be used to create clones of genes, plasmids are called cloning vectors.
3. Getting the Plasmid In
In nature bacteria have various enzymes that cut up the DNA of their natural enemies, such as bacteriophages (bacterial viruses). Researchers have taken advantage of these so-called restriction enzymes to splice DNA for use in engineering bacteria. Hundreds of restriction enzymes have been isolated and each will cut a DNA strand at a specific sequence of nucleotides. Some restriction enzymes generate blunt ends, cutting across both strands of DNA. Others generate a staggered cut, producing “sticky ends.” These ends anneal by hydrogen bonding to similar ends on another DNA segment cut with the same restriction enzyme.
Cloning a gene involves identifying a gene of interest in an organism, isolating DNA from that organism, and then using a restriction enzyme to snip the gene from the DNA strand. The gene?containing segment can then be spliced into a plasmid cut by the same restriction enzyme. The bacteria take up the plasmid and are allowed to replicate.
Ordinarily, bacterial cells do not readily take up plasmids. Researchers can use various tricks, however, to get cells more ready to do so. One common method holds the cells on ice in a solution of calcium chloride. The cells are then briefly heat shocked so the plasmid can cross the plasma membrane. An alternate method,electroporation">electroporation, uses a short electrical pulse to open pores in the plasma membrane, allowing the plasmid to pass through.
Marker genes, such as genes for antibiotic resistance, are often engineered into plasmids. These marker genes enable researchers to know which bacteria have the plasmids. The antibiotic is added to the media used to grow the bacteria. Cells that do not contain the plasmid will fail to reproduce. In addition to marker genes, plasmids typically contain one or more genes of interest. For example, a protein not otherwise expressed by the recipient cell might be produced only when the plasmid is present. Individual colonies of bacteria, each derived from a single cell, can be evaluated for the expression of such novel gene products.
Protein production can be straightforward if the source of the novel gene was another bacterium. However, the goal of modifying bacteria might be the production of proteins encoded by eukaryotic genes from fungi, plants or animals. This presents challenges. Eukaryotic DNA contains both exons">exons (coding sequences) and introns">introns (intervening sequences). In eukaryotic cells this DNA is used as a template for the production of mRNA, which must then undergomRNA splicing">mRNA splicing. Introns are removed and exons are joined to form the mRNA, which travels to the ribosome for protein production. Bacteria lack the enzymes necessary for mRNA splicing, so introducing a eukaryotic gene into bacteria requires a special procedure. First, DNA must be generated that is complementary to the already spliced mRNA. The enzymereverse transcriptase">reverse transcriptase is then used to generate a double-stranded DNA molecule called cDNA, using the mRNA as a template. Finally, this cDNA is incorporated into the cloning vector.
Expressing eukaryotic genes in bacteria presents other problems. After proteins are assembled in eukaryotic cells they are often modified. (See the Proteins and Proteomics unit.) For example, various sugars may be attached to the polypeptide so that glycoproteins are formed. Bacteria are generally unable to accomplish such post-translational modifications, and eukaryotic genes expressed in bacteria may not function properly. The inability of bacteria to perform such modifications has driven scientists to use yeast (Saccharomyces cerevisiae) and eukaryotic cell culture to produce some recombinant products.
4. Are Recombinant Bacteria Safe?
Concerns about the safety of recombinant bacteria were voiced as the technology was developed. Some fear that new, untreatable human pathogens could be inadvertently generated. In 1974 prominent researchers self-imposed a moratorium on certain experiments until they could assess the hazards. After much discussion, the researchers developed biological containment procedures. These include generating recombinant DNA only in bacteria that have mutations to prevent them from surviving outside of the laboratory. The release of recombinant microbes into the environment remains controversial.
5. Genetic Modification of Plants
New traits introduced to crop plants by genetic engineering have the potential to increase crop yields, improve agricultural practices, or add nutritional quality to products. For example, transgenic crop plants capable of degrading weed killers allow farmers to spray weeds without affecting yield. Use of herbicide-tolerant crops may also allow farmers to move away from pre-emergent herbicides and reduce tillage, thereby decreasing soil erosion and water loss. Transgenic plants that express insecticidal toxins resist attacks from insects. Crops engineered to resist insects are an alternative to sprays, which may not reach all parts of the plant. They are also cost effective, reducing the use of synthetic insecticides. Genetic engineering has also been used to increase the nutritional value of food; “golden rice” is engineered to produce beta-carotene, for example. Edible vaccines, present in the plants we eat, may be on the horizon.
The new traits expressed in such transgenic plants are derived from a variety of other organisms. Scientists have given a gene from the bacterium Salmonella to cultivars of soybeans, corn, canola, and cotton to degrade the pesticide glyphosphate (Roundup TM). The gene for the insecticidal toxin in transgenic cotton, potato, and corn plants comes from the bacterium Bacillus thuringiensis(Bt). One of the genes allowing vitamin A production in golden rice is derived from the bacterium Erwinia uredovora; others are from the daffodil.

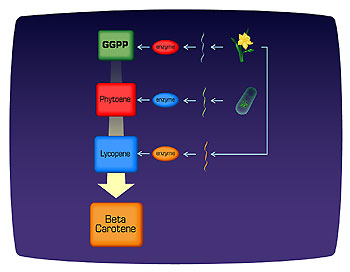
Fig 2. Biochemical pathway for beta-carotene production in golden rice.
The biochemical pathway for beta-carotene synthesis in golden rice.
The development of golden rice involved the introduction of several genes into a plant to provide a multistep biochemical pathway
(Fig 2). Rice grain, which serves as a food staple for much of the world, lacks vitamin A. An estimated 100 million to 200 million children worldwide have vitamin A deficiency, a condition that causes blindness; and increases susceptibility to diarrhea, respiratory infection, and childhood diseases, such as measles. Beta-carotene and other carotenes (the red, yellow, and orange pigments found in carrots and other vegetables) are the precursors of vitamin A. Rice synthesizes beta-carotene in its chloroplasts but not in the edible seed tissue.
Ingo Potrykus and his colleagues found that geranyl geranyl diphosphate (GGPP), a precursor to carotenoid production, is present in rice seed. They genetically engineered golden rice to express the enzymes necessary for the conversion of GGPP to beta-carotene. The synthesis of beta-carotene from geranyl geranyl diphosphate requires four biochemical reactions, each catalyzed by a different enzyme. A bacterium, Agrobacterium tumefaciens, containing three plasmids, was used to introduce all the genes necessary for the complete biochemical pathway for beta-carotene production. It was possible to use three enzymes instead of four because the bacterial enzyme phytoene desaturase accomplishes what two plant enzymes (phytoene desaturase and beta-carotene desaturase) do.
If transgenic plants can help prevent vitamin deficiencies, can they also produce vaccines? Edible vaccines available in crops could help people in developing nations where transportation, refrigeration, and disposable needle supplies are limited. Hugh Mason and his colleagues (Boyce Thompson Institute) have expressed a gene that encodes an E. coli protein in potatoes. Volunteers who ate raw, modified potatoes developed antibodies to the protein. Research is underway to see whether the antibodies will protect against diarrhea induced by disease-causing E. coli.
6. Techniques Used for Generating Transgenic Plants
As with bacteria, the ability to genetically modify plants depends on obtaining genetically identical populations and readily manipulating DNA. How do you “clone” a plant? Many plant species naturally undergo asexual reproduction by fragmentation, where segments from a parent plant regenerate a new plant. It is also possible to grow plants in culture from small explants. Another method is to culture plants from totipotent">totipotent cells found in plant meristems. These plant cells can divide and differentiate into the various types of specialized cells. In a test tube, plant cells will divide and form an undifferentiated callus. When hormones in the culture medium are adjusted, the callus will sprout shoots and roots and eventually develop into a plantlet that can be transplanted to soil. To clone a plant – perhaps a plant with new genes – the growing callus is simply subdivided. Thousands of genetically identical plants can be generated in this way.
How do you get a plant to take up a gene? Researchers working with rice often use the soil bacterium Agrobacterium tumefaciens. This bacterium, the cause of crown gall disease in many fruit plants, is well known for its ability to infect plants with a tumor-inducing (Ti) plasmid. A section of the Ti plasmid, called T-DNA, integrates into chromosomes of the plant. Recombinant DNA can be added to the T-DNA, the gall-inducing genes removed, and infection by the bacteria – containing the recombinant plasmid – will provide for transfer of novel genes to plant embryos.

Figure 3. A “gene gun”
GENE GUN. Courtesy of Bio-Rad Laboratories.
Although Agrobacterium tumefaciens works for introducing plasmids into rice, not all plants are equally susceptible to this bacterium. Researchers interested in modifying crops such as wheat and corn have turned to other methods for delivering genes to plant cells. One approach is to use a “gene gun">gene gun,” (Fig. 3) which fires plastic bullets filled with DNA-coated metallic pellets. An explosive blast or burst of gas propels the bullet toward a stop plate. The DNA-coated pellets are directed through an aperture in the stop plate, and then penetrate the walls and membranes of their cellular targets. Some projectiles penetrate the nuclei of cells, where occasionally the introduced DNA integrates into the DNA of the plant genome. Transformed cells can then be cloned in culture.
Marker genes are often included in DNA constructs so that plants that have acquired the novel DNA can be selected. In plants, marker genes include those for herbicide resistance. Plants that grow in the presence of the herbicide are assumed to possess the transgene of interest. The transgenic plant embryos are cultivated in tissue culture. Once mature plants are obtained they are evaluated for the activity of the introduced gene, any unintended effect on plant growth, and product yield and quality. The ability of the gene to be expressed in subsequent plant generations is also evaluated.
Not all genes are expressed in every tissue of a plant. When golden rice was developed it was necessary to ensure that the novel genes were expressed in the endosperm of the seed. The endosperm of a seed is the starchy component that provides energy and nutrients for the developing plant embryo. Regulatory DNA sequences upstream from the specified genes were introduced into the recombinant Ti plasmids. Such regulatory regions influence where and when a gene will be expressed. (See the Genomics unit.) The regulatory regions chosen for golden rice provide an uninhibited transcription of the genes in endosperm.
7. Problems and Concerns
Several concerns have been raised regarding transgenic crop plants. Foremost is the possibility that the process of genetic engineering might inadvertently generate new allergens or toxins that could affect human health. Another concern is that introduced genes from engineered crops could move to other organisms in the environment. Other concerns relate to cultivars that are engineered to produce insecticides. The potential development of insecticide resistance in target pests is worrisome; as is the possibility that non-target, beneficial insects might be affected by engineered plants.
A particular concern is the possibility that transgenic crop plants could affect human health by expressing unanticipated allergens. In March 1996 researchers at the University of Nebraska showed that an allergen from Brazil nuts had been transferred into soybeans. Individuals sensitized to Brazil nuts make antibodies (IgE) specific to certain proteins in the nuts. Engineered soybeans reacted with such antibodies in vitro. Had allergic individuals consumed the transgenic soybeans they would have likely experienced IgE-mediated reactions, ranging from itching to anaphylaxis.
Obviously, expressing a known allergen in food crops is unwise. However, it is difficult to predict whether a protein expressed in a novel organism will cause allergies. A protein isolated from its native species may differ from the same protein (with an identical amino acid sequence) harvested from a transgenic organism expressing that protein. Sometimes sugar or acetyl groups are added to proteins after they are manufactured at the ribosome. The forms of sugar or acetyl groups may vary between organisms. Sugar groups on proteins have been associated with allergenic and immunogenic responses. Hence, allergenicity studies ought to be carried out on the actual material derived from transgenic plants themselves, rather than on just the bacterial proteins. Such studies are not always done.
Critics are worried that engineered plants might generate toxins as a result of the DNA-insertion process. They note that the insertion of genetic material (using gene gun technology, for example) is semi-random, and that the amount and location of DNA inserted into the chromosome varies. If an insert disrupts a regulatory region that serves to “turn off” the production of a toxin, the result might be an over-expression of toxin. Another concern is the inclusion of regulatory regions as part of genetic constructs: the regulation of host genes near an insert could be dramatically affected.
Significant concerns relate to the impact that genetically modified plants could make on the environment. In experiments, transgenic crops are known to hybridize with closely related species. The probability that transgenic traits, as well as other accompanying changes in traits, will show up in wild plant relatives is increasing as genetically modified crops are established. Herbicide-tolerant weeds can evolve; glyphosphate- (Roundup TM) resistant rigid ryegrass, for example, has developed only recently. Genetically modified crops must be monitored to reduce unintended degradation of natural ecosystems.
Crops engineered to produce insecticides, such as Bt toxin, bring other concerns. The widespread planting of Bt corn and other crops can result in insects evolving a resistance to Bt toxins. At least ten species of moths, two species of beetles, and four species of flies already have developed resistance to Bt toxins under laboratory exposure. Bt toxins administered as a spray are present only transiently. However, transgenic crops continuously express the insecticidal protein. This ongoing exposure may be more likely to select for resistant insects.
The emergence of a resistant insect population is likely whenever a pesticide is used. One strategy for delaying the emergence of insects resistant to Bt toxin is to plant a “refuge” of conventional crops near Bt-expressing crops. The idea is that these conventional crops will harbor susceptible insects that will mate with resistant insects, diluting out recessive resistance alleles. Of course, if resistance develops as a dominant allele, this strategy will not work.
There are hundreds of known subspecies of Bacillus thuringiensis, and the insecticidal toxin derived from each is poisonous only to certain species of insects. Nevertheless, there are concerns that plants expressing genes for such toxins could affect non-target insect species. Some of these species may be beneficial, such as those that provide pollination or consume pests. Laboratory experiments suggest an increased mortality of Monarch butterflies that ingested Bt corn pollen. How frequently this occurs in the field is unknown, and not all laboratory studies have given similar results. The Environmental Protection Agency (EPA) requires toxicity tests on a standard set of organisms before a pesticide can be registered. As of December 2002 the EPA had not demonstrated toxicity of Bt to nontarget species. Data gathering continues.
8. Genetic Modification of Animals
Dolly the lamb stole the headlines as the first example of livestock cloned from DNA of an adult animal. But the real breakthrough came with Polly, the first transgenic lamb. Born the year after Dolly, Polly was given a human gene that encodes blood-clotting factor IX, the protein missing in people with one form of hemophilia. Harvesting such proteins from transgenic livestock is one goal of this research. The road to Polly and subsequent transgenic animals began with research using genetically altered mice. Along the way, technologies for cloning animals, modifying DNA, and targeting expression of proteins to specific tissues were developed. Someday, human gene therapy – supplying genes to patients with missing or altered proteins – may become common practice. However, significant challenges remain. Moreover, risks and ethical concerns must be addressed.
Antithrombin III (AT-III) is an example of a pharmaceutical produced in transgenic livestock. A normal level of AT-III keeps the formation of blood clots under control. Patients with AT-III deficiency may have thromboembolic problems beginning in early adulthood, particularly clots in the legs and pulmonary embolism. Providing therapeutic AT-III can reduce clotting risks in such patients. Other therapeutic proteins being considered for production by transgenic animals include human hemoglobin, human serum albumin, tissue plasminogen activator (used to treat stroke), human alpha-1-antitrypsin (alpha-1-antitrypsin deficiency can cause life-threatening emphysema), various vaccine proteins, and monoclonal antibodies.
For some time, mice have been genetically altered to exhibit human genetic disease. To generate such animal models normal genes in mice are inactivated using “knockout” technology, or altered by replacement of the normal gene with a mutated counterpart. Mouse disease models now exist for cystic fibrosis, beta-thalassaemia, atherosclerosis, retinoblastoma, and Duchenne muscular dystrophy. Such animal models allow researchers to test therapeutic compounds and study the molecular basis of given diseases.

Figure 4. Gene disruption by homologous recombination
The plasmid contains a gene interrupted by a marker gene (XR). Recombination involving two crossovers between the plasmid and wild type chromosomal DNA with the interrupted gene and the selectable marker.
Knockout technology, as well as other genetic engineering approaches, depends on the ability to target genes for insertion into particular locations within the host chromosome. To do this, a region on the chromosome is identified and DNA homologous to that region is engineered into a cloning vector. The newly inserted sequence can then be disrupted by insertion of a selected gene; for example, a marker gene encoding antibiotic resistance. Once cells take up the DNA, homologous recombination on either side of the marker gene allows it to be precisely inserted into the chromosome. At the same time, some or all of the target gene on the chromosome is deleted (Fig. 4).
Gene knockout in pigs is being studied as an avenue for transplanting animal organs into humans. A major cause of tissue rejection is an immune reaction to the carbohydrate galactose-a-1,3,-galactose on the surface of non-human cells. Deletion of the a-1,3,-galatosyltransferase gene may allow the production of animals lacking this surface marker.
As researchers recognized the potential of transgenic livestock for the production of human therapeutics and transplant tissue, farmers recognized the contributions that genetic engineering might make to the economics of livestock production. Cows might be produced that could grow more muscle mass, require less feed, produce more milk, or be leaner. The composition of milk could be changed; for example, casein could be over-expressed to provide increased cheese production. Lactose might be removed from milk for lactose-intolerant consumers. Disease resistant animals could reduce the use of antibiotics. Poultry with less fat content and eggs with lower cholesterol are other goals.
9. Cloning Animals
Asexual reproduction in bacteria and plants allows scientists to obtain genetically identical populations; this does not occur naturally in vertebrates, except in twins. In 1996 Dolly the lamb was born: chromosomal material derived from an adult sheep was used to generate an animal with chromosomal DNA identical to that of the donor animal. Cloning livestock, using the techniques that generated Dolly, may become an economical method for traditional breeders to replicate their superior animals and provide them to farmers. Rather than selling semen, breeding companies might distribute cloned embryos for implantation into surrogate cows. Because Dolly did not possess foreign DNA, she was not transgenic. However, she did represent a valuable step toward the development of transgenic livestock. With donor DNA for cloning derived from cultured recombinant cells, it becomes possible to carry out specific genetic modifications and introduce the modified genes into animals.
Figure 5. A donor cell is fused with enucleated egg cell by subjecting the two cells to pulses of electricity. The cell replicates in culture, generating an embryo, which is then introduced into the uterus of a female for development.
Nuclear Transfer
Ian Wilmut and his colleagues cloned Dolly using a technique called nuclear transfer 1. In this technique, the nucleus of a recipient egg is removed to make way for the genetic material of the donor (Fig. 5). The donor cell is fused with the enucleated egg cell by subjecting the two cells to pulses of electricity. Earlier studies had suggested that donor nuclei from early embryos were more likely develop properly. The use of an adult cell for the donor nucleus was unique in Dolly’s case. Although most differentiated animal cells contain all the genes for making an entire organism, nuclei change as cells differentiate. To dedifferentiate the udder cells used for nuclear transfer they cultured the cells in a nutrient-poor medium. This caused the cell cycle to stop in the GO phase. After fusion, 277 embryos were grown in culture for six days before implanting them in thirteen surrogate mothers. Only one of the embryos completed normal development.
Cloning by nuclear transfer depends on the availability of donor cells with the appropriate genetic information. Somatic cells such as fibroblasts, ovarian cells, muscle cells, and mammary epithelia are grown in cell culture and by genetically modified fusion with the enucleated egg. Commonly, DNA is transferred to the cells using viruses.

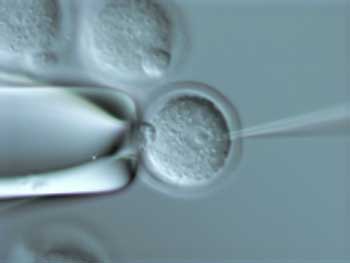
Figure 6: Microinjection
Photo of micro-injection.
Anne Bower and Manfred Baetsher, MICROINJECTION. Courtesy of Oregon Health and Science University, Transgenic/Gene Targeting Facility.
Microinjection and Other Techniques
Another technique for cloning animals is microinjection. In this technique, a gene construct is characterized in culture and an adequate quantity of the desired DNA is obtained. The DNA is injected into fertilized ova before the first cell division occurs. This increases the probability that all of the cells of the organisms will harbor the gene. The injection is done soon after fertilization, before the male and female pronuclei have fused. A very thin pipette or needle injects the DNA into the large male pronucleus (Fig. 6). Surrogate mothers are made pseudo-pregnant with hormones and implanted with the injected eggs. After birth, tissue samples of the young are assessed for the presence of the desired gene. DNA from germ line cells is given special attention. If the novel gene is present in these cells, the animal can be used as a founder for breeding.
Genetic constructs that include regulatory regions targeting gene expression to specific tissues are necessary if the gene product is to be harvested readily. For example, GTC Biotherapeutics uses the betacasein promoter to ensure that antithrombin III is secreted in goat milk. Common biochemical procedures, such as filtration and chromatography, are then used to isolate the AT-III from the milk.

Figure 7. Southern blotting
1) DNA fragments are generated using restriction enzymes. 2) The fragments are separated in a gel by the application of an electric charge. 3) The fragments are then blotted onto a piece of nitrocellulose paper, where they retain their same pattern of separation, but are denatured to become single-stranded DNA. 4) A unique single-stranded portion of the gene of interest (the probe) is radioactively labeled and allowed to anneal with the blotted paper. 5) When exposed to a sheet of photographic film, any DNA fragments that annealed with the labeled probe are identified.
Scientists often use southern blots to evaluate DNA extracts from tissue samples (Fig. 7). Southern blotting a type of nucleic acid hybridization test, in which single-stranded DNA from two sources interact. Strands with similar nucleic acid sequences will anneal by base pairing (A with T, and G with C) to form double-stranded molecules. One of the single-stranded DNA molecules is a unique portion of the gene of interest, and is radiolabeled so it can be detected on photographic film (the probe). Southern blotting allows the detection of fragments of genomic DNA, which anneal to the radiolabeled probe. The fragments are generated using restriction enzymes and separated in a gel by electrophoresis. The size of a given fragment relates to the distance it migrates on electrophoresis. The fragments are denatured to single strands, transferred to a special filter paper that is immersed in a solution containing the probe, and then rinsed. If the probe has annealed it will expose the photographic film, resulting in a band.
10. Challenges
Even beyond the controversies involving human cloning, there are risks and ethical dilemmas surrounding the use of transgenic and cloned animals.
One risk from cloning animals is a loss of genetic diversity in livestock. This could result in increased susceptibility to disease or other environmental challenges. Some of this risk might be avoided, according to the Roslin Institute, by systems that limit the number of clones produced by breeders and restricting the number of clones sold to any given farmer.
The overexpression or deletion of certain genes must also be evaluated from an animal welfare perspective. The secretion of proteins in the milk of transgenic goats seems to have no ill effects. However, pigs that harbor foreign genes have exhibited many problems including lameness, lethargy, thickened skin, kidney dysfunction, inflamed joints, peptic ulcers, pericarditis, severe osteoarthritis, and a propensity toward pneumonia.
The safety of cloning techniques has been questioned by a number of researchers. Rudolf Jaenisch (MIT) published a study in September 2002 comparing 10,000 genes from placentas and livers of newborn cloned mice with those from normal mice; at least four percent were functioning incorrectly. Cloned mice have exhibited developmental abnormalities, obesity, pneumonia, liver failure, and premature death. Dolly exhibited arthritis at an unusually young age and was put to sleep at age six, about half the life expectancy of sheep in captivity.
An additional concern about the use of transgenic animal products, including transplanted organs, is the risk of human exposure to animal pathogens. At least 150 pathogens are known to infect both humans and some other animal. In 1997 the isolation of two retroviruses from pigs that could infect human tissue culture cells was reported. These so called PERVs (porcine endogenous retroviruses) are of special concern to those considering the use of porcine tissue for transplants, especially because some retroviruses have been associated with cancer.
11. Addressing the Controversies
Decisions made regarding the use of genetically modified organisms will impact the environment, and force a reexamination of consumer safety and animal welfare issues. Do the benefits provided by transgenic organisms outweigh the risks? Are those making decisions influenced too heavily by the profit motive? How can opportunities for competing approaches be ensured?
Certainly the production of genetically altered organisms is a profit-making business. In 1980, individuals and companies realizing this sought protection of their intellectual property and turned to the courts. That year the U.S. Supreme Court delivered a landmark decision stating that living organisms are patentable; in 1988 a patent was issued for the genetically altered “Harvard mouse.”
In late 2001 seventy-seven scientists and teachers from sixteen countries, concerned with how environmental protection decisions are made, issued the Lowell Statement on Science and Precaution. Their “Precautionary Principle” recommends using the safest approaches to meeting society’s needs, placing responsibility for finding the safest alternatives in the hands of those originating potentially dangerous activities, use of independent review, and participation of those who may be affected by a policy choice. These guidelines might well be extended beyond environmental policy.
Governmental bodies often play the role of reviewer when it comes to safety, particularly of foods. Various governments and organizations have begun generating guidelines and recommendations regarding foods derived from transgenic organisms. For example, the Food and Agricultural Organization of the United Nations along with the World Health Organization organized a series of scientific consultations to provide their member nations with recommendations. In a January 2001 report the consultation agreed that “the safety assessment of foods derived from biotechnology requires an integrated and stepwise, case-by-case approach.” 2
Can the population at large – by consumer and political choice – influence the use of genetically modified organisms? In November 2002 Oregon was the first state in the U.S. to put labeling of genetically modified foods on the ballot. Proponents of labeling spent about 200,000 dollars to convince voters. Opponents, with funding from large agribusinesses, spent 5.5 million dollars to kill the idea. Voters were convinced that labeling would significantly increase food costs and rejected the measure.
End Notes
-
- Wilmut I., A. E. Schnieke, J. McWhir, A. J. Kind, and K. H. S. Canbell. 1997. Viable offspring derived from fetal and adult mammalian cells. Nature 385:810-12.
- Food and Agriculture Organization of the United Nations (FAO). 2001. Evaluation of Allergenicity of Genetically Modified Foods. Report of a Joint FAO/WHO Expert Consultation on Allergenicity of Food Derived from Biotechnology 22-25 Jan 2001. Rome, Italy