Rediscovering Biology: Molecular to Global Perspectives
Online Textbook and Video
This online textbook chapter supports and extends the content of the Cell Biology and Cancer video. The chapter covers cancer causes and treatments, and tumor biology and angiogenesis. Click on the Go button to begin reading, or skip to a sub-section of the chapter in the list below.
1. Introduction
“We now understand a lot about cancer. We know that it results from a series of genetic changes having to do with cell division and growth control and genetic instability, mortality, the suicide mechanism in cells; the ability of the cells to migrate; the ability of the cells to attract to them a blood supply. And so that’s pretty profound that in a few sentences one can summarize a sophisticated, fundamental understanding of what a cancer is.”
— Leland Hartwell
A multicellular organism can thrive only when all its cells function in accordance with the rules that govern cell growth and reproduction. Why does a normal cell suddenly become a “rebel,” breaking the rules, dividing recklessly, invading other tissues, usurping resources, and in some cases eventually killing the body in which it lives?
To understand how and why cells rebel, we need to understand the normal functions of cell growth and reproduction. From the mid-nineteenth century on, research in cell biology, biochemistry, and molecular biology has provided astonishingly detailed information about the molecules and processes that allow cells to divide, grow, differentiate, and perform their essential functions. This basic knowledge of cell biology has also led to practical discoveries about the mechanisms of cancer. Specific molecules that control the progression of a cell through the cell cycle regulate cell growth. An understanding of normal cell cycle processes and how those processes go awry provides key information about the mechanisms that trigger cancer. Loss of control of the cell cycle is one of the critical steps in the development of cancer.
Although cancer comprises at least 100 different diseases, all cancer cells share one important characteristic: they are abnormal cells in which the processes regulating normal cell division are disrupted. That is, cancer develops from changes that cause normal cells to acquire abnormal functions. These changes are often the result of inherited mutations or are induced by environmental factors such as UV light, X-rays, chemicals, tobacco products, and viruses. All evidence suggests that most cancers are not the result of one single event or factor. Rather, around four to seven events are usually required for a normal cell to evolve through a series of premalignant stages into an invasive cancer. Often many years elapse between the initial event and the development of cancer. The development of molecular biological techniques may help in the diagnosis of potential cancers in the early stages, long before tumors are visible.
2. What Is Cancer?
Cancer results from a series of molecular events that fundamentally alter the normal properties of cells. In cancer cells the normal control systems that prevent cell overgrowth and the invasion of other tissues are disabled. These altered cells divide and grow in the presence of signals that normally inhibit cell growth; therefore, they no longer require special signals to induce cell growth and division. As these cells grow they develop new characteristics, including changes in cell structure, decreased cell adhesion, and production of new enzymes. These heritable changes allow the cell and its progeny to divide and grow, even in the presence of normal cells that typically inhibit the growth of nearby cells. Such changes allow the cancer cells to spread and invade other tissues.
The abnormalities in cancer cells usually result from mutations in protein-encoding genes that regulate cell division. Over time more genes become mutated. This is often because the genes that make the proteins that normally repair DNA damage are themselves not functioning normally because they are also mutated. Consequently, mutations begin to increase in the cell, causing further abnormalities in that cell and the daughter cells. Some of these mutated cells die, but other alterations may give the abnormal cell a selective advantage that allows it to multiply much more rapidly than the normal cells. This enhanced growth describes most cancer cells, which have gained functions repressed in the normal, healthy cells. As long as these cells remain in their original location, they are considered benign; if they become invasive, they are considered malignant. Cancer cells in malignant tumors can often metastasize, sending cancer cells to distant sites in the body where new tumors may form.
3. Genetics of Cancer
Only a small number of the approximately 35,000 genes in the human genome have been associated with cancer. (See the Genomics unit.) Alterations in the same gene often are associated with different forms of cancer. These malfunctioning genes can be broadly classified into three groups. The first group, called proto-oncogenes, produces protein products that normally enhance cell division or inhibit normal cell death. The mutated forms of these genes are called oncogenes. The second group, called tumor suppressors, makes proteins that normally prevent cell division or cause cell death. The third group contains DNA repair genes, which help prevent mutations that lead to cancer.
Proto-oncogenes and tumor suppressor genes work much like the accelerator and brakes of a car, respectively. The normal speed of a car can be maintained by controlled use of both the accelerator and the brake. Similarly, controlled cell growth is maintained by regulation of proto-oncogenes, which accelerate growth, and tumor suppressor genes, which slow cell growth. Mutations that produce oncogenes accelerate growth while those that affect tumor suppressors prevent the normal inhibition of growth. In either case, uncontrolled cell growth occurs.
Oncogenes and Signal Transduction
In normal cells, proto-oncogenes code for the proteins that send a signal to the nucleus to stimulate cell division. These signaling proteins act in a series of steps called signal transduction cascade or pathway (Fig. 1).
Figure 1. A signal (in this example, a growth factor) binds to a tyrosine kinase receptor on the outside of the cell. This activates the membrane protein (through the addition of phosphate groups), which in turn activates proteins in the cytoplasm such as kinases. Several other proteins may be involved in the cascade, ultimately activating one or more transcription factors. The activated transcription factors enter the nucleus, where they stimulate the expression of the genes that are under the control of that factor. This is an example of the Ras pathway, which results in cell division.
(See the Genetics and Development unit.) This cascade includes a membrane receptor for the signal molecule, intermediary proteins that carry the signal through the cytoplasm, and transcription factors in the nucleus that activate the genes for cell division. In each step of the pathway, one factor or protein activates the next; however, some factors can activate more than one protein in the cell. Oncogenes are altered versions of the proto-oncogenes that code for these signaling molecules. The oncogenes activate the signaling cascade continuously, resulting in an increased production of factors that stimulate growth. For instance, MYC is a proto-oncogene that codes for a transcription factor. Mutations in MYC convert it into an oncogene associated with seventy percent of cancers. RAS is another oncogene that normally functions as an “on-off” switch in the signal cascade. Mutations in RAS cause the signaling pathway to remain “on,” leading to uncontrolled cell growth. About thirty percent of tumors — including lung, colon, thyroid, and pancreatic carcinomas — have a mutation in RAS.
The conversion of a proto-oncogene to an oncogene may occur by mutation of the proto-oncogene, by rearrangement of genes in the chromosome that moves the proto-oncogene to a new location, or by an increase in the number of copies of the normal proto-oncogene. Sometimes a virus inserts its DNA in or near the proto-oncogene, causing it to become an oncogene. The result of any of these events is an altered form of the gene, which contributes to cancer. Think again of the analogy of the accelerator: mutations that convert proto-oncogenes into oncogenes result in an accelerator stuck to the floor, producing uncontrolled cell growth.
Most oncogenes are dominant mutations; a single copy of this gene is sufficient for expression of the growth trait. This is also a “gain of function” mutation because the cells with the mutant form of the protein have gained a new function not present in cells with the normal gene. If your car had two accelerators and one was stuck to the floor, the car would still go too fast, even if there were a second, perfectly functional accelerator. Similarly, one copy of an oncogene is sufficient to cause alterations in cell growth. The presence of an oncogene in a germline cell (egg or sperm) results in an inherited predisposition for tumors in the offspring. However, a single oncogene is not usually sufficient to cause cancer, so the inheritance of an oncogene does not necessarily result in cancer.
Tumor Suppressor Genes
The proteins made by tumor suppressor genes normally inhibit cell growth, preventing tumor formation. Mutations in these genes result in cells that no longer show normal inhibition of cell growth and division. The products of tumor suppressor genes may act at the cell membrane, in the cytoplasm, or in the nucleus. Mutations in these genes result in a loss of function (that is, the ability to inhibit cell growth) so they are usually recessive. This means that the trait is not expressed unless both copies of the normal gene are mutated. Using the analogy to a car, a mutation in a tumor suppressor gene acts much like a defective brake: if your car had two brakes and only one was defective, you could still stop the car.
How is it that both genes can become mutated? In some cases, the first mutation is already present in a germline cell (egg or sperm); thus, all the cells in the individual inherit it. Because the mutation is recessive, the trait is not expressed. Later a mutation occurs in the second copy of the gene in a somatic cell. In that cell, both copies of the gene are mutated and the cell develops uncontrolled growth. An example of this is hereditary retinoblastoma, a serious cancer of the retina that occurs in early childhood. When one parent carries a mutation in one copy of the RB tumor suppressor gene, it is transmitted to offspring with a fifty percent probability. About ninety percent of the offspring who receive the one mutated RB gene from a parent also develop a mutation in the second copy of RB, usually very early in life. These individuals then develop retinoblastoma. Not all cases of retinoblastoma are hereditary: it can also occur by mutation of both copies of RB in the somatic cell of the individual. Because retinoblasts are rapidly dividing cells and there are thousands of them, there is a high incidence of a mutation in the second copy of RB in individuals who inherited one mutated copy. This disease afflicts only young children because only individuals younger than about eight years old have retinoblasts. In adults, however, mutations in RB may lead to a predisposition to several other forms of cancer.
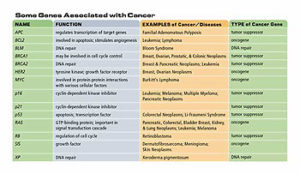
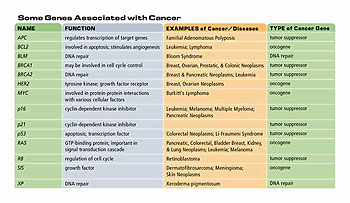
Table 1. Some genes associated with cancer
Three other cancers associated with defects in tumor suppressor genes include familial adenomatous polyposis of the colon (FPC), which results from mutations to both copies of the APC gene; hereditary breast cancer, resulting from mutations to both copies of BRCA2; and hereditary breast and ovarian cancer, resulting from mutations to both copies of BRCA1. While these examples suggest that heredity is an important factor in cancer, the majority of cancers are sporadic with no indication of a hereditary component. Cancers involving tumor suppressor genes are often hereditary because a parent may provide a germline mutation in one copy of the gene. This may lead to a higher frequency of loss of both genes in the individual who inherits the mutated copy than in the general population. However, mutations in both copies of a tumor suppressor gene can occur in a somatic cell, so these cancers are not always hereditary. Somatic mutations that lead to loss of function of one or both copies of a tumor suppressor gene may be caused by environmental factors, so even these familial cancers may have an environmental component.
DNA Repair Genes
A third type of gene associated with cancer is the group involved in DNA repair and maintenance of chromosome structure. Environmental factors, such as ionizing radiation, UV light, and chemicals, can damage DNA. Errors in DNA replication can also lead to mutations. Certain gene products repair damage to chromosomes, thereby minimizing mutations in the cell. When a DNA repair gene is mutated its product is no longer made, preventing DNA repair and allowing further mutations to accumulate in the cell. These mutations can increase the frequency of cancerous changes in a cell. A defect in a DNA repair gene called XP (Xeroderma pigmentosum) results in individuals who are very sensitive to UV light and have a thousand-fold increase in the incidence of all types of skin cancer. There are seven XP genes, whose products remove DNA damage caused by UV light and other carcinogens. Another example of a disease that is associated with loss of DNA repair is Bloom syndrome, an inherited disorder that leads to an increased risk of cancer, lung disease, and diabetes. The mutated gene in Bloom syndrome, BLM, is required for maintaining the stable structure of chromosomes. Individuals with Bloom syndrome have a high frequency of chromosome breaks and interchanges, which can result in the activation of oncogenes.
4. Cell Cycle
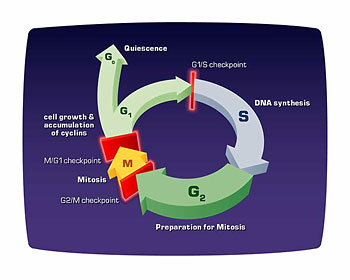
Normal cells grow and divide in an orderly fashion, in accordance with the cell cycle. (Mutations in proto-oncogenes or in tumor suppressor genes allow a cancerous cell to grow and divide without the normal controls imposed by the cell cycle.) The major events in the cell cycle are described in Fig. 2.
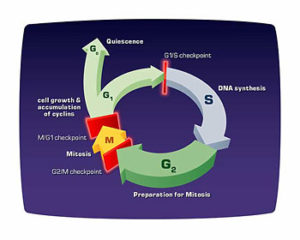
Figure 2. The cell cycle is an ordered process of events that occurs in four stages. During the two gap phases, G1 and G2, the cell is actively metabolizing but not dividing. In S (synthesis) phase, the chromosomes duplicate as a result of DNA replication. During the M (mitosis) phase the chromosomes separate in the nucleus and the division of the cytoplasm (cytokinesis) occurs. There are checkpoints in the cycle at the end of G1 and G2 that can prevent the cell form entering the S or M phases of the cycle. Cells that are not in the process of dividing are in the G0 stage, which includes most adult cells.
Several proteins control the timing of the events in the cell cycle, which is tightly regulated to ensure that cells divide only when necessary. The loss of this regulation is the hallmark of cancer. Major control switches of the cell cycle are cyclin-dependent kinases. Each cyclin-dependent kinase forms a complex with particular cyclins, a protein that binds and activates the cyclin-dependent kinase. The kinase part of the complex is an enzyme that adds a phosphate to various proteins required for the progression of a cell through the cycle. These added phosphates alter the structure of the protein and can activate or inactivate the protein, depending on its function. There are specific cyclin-dependent kinase/cyclin complexes at the entry points into the G1, S, and M phases of the cell cycle, as well as additional factors that help prepare the cell to enter S phase and M phase.
One important protein in the cell cycle is p53, a transcription factor (see Genetics of Development unit) that binds to DNA, activating transcription of a protein called p21. P21 blocks the activity of a cyclin-dependent kinase required for progression through G1. This block allows time for the cell to repair the DNA before it is replicated. If the DNA damage is so extensive that it cannot be repaired, p53 triggers the cell to commit suicide. The most common mutation leading to cancer is in the gene that makes p53.
Li-Fraumeni syndrome, an inherited predisposition to multiple cancers, results from a germline (egg or sperm) mutation in p53. Other proteins that stop the cell cycle by inhibiting cyclin-dependent kinases are p16 and RB. All of these proteins, including p53, are tumor suppressors.
Cancer cells do not stop dividing, so what stops a normal cell from dividing? In terms of cell division, normal cells differ from cancer cells in at least four ways.
-
- Normal cells require external growth factors to divide. When synthesis of these growth factors is inhibited by normal cell regulation, the cells stop dividing. Cancer cells have lost the need for positive growth factors, so they divide whether or not these factors are present. Consequently, they do not behave as part of the tissue – they have become independent cells.
-
- Normal cells show contact inhibition; that is, they respond to contact with other cells by ceasing cell division. Therefore, cells can divide to fill in a gap, but they stop dividing as soon as there are enough cells to fill the gap. This characteristic is lost in cancer cells, which continue to grow after they touch other cells, causing a large mass of cells to form.
-
- Normal cells age and die, and are replaced in a controlled and orderly manner by new cells. Apoptosis is the normal, programmed death of cells. Normal cells can divide only about fifty times before they die. This is related to their ability to replicate DNA only a limited number of times. Each time the chromosome replicates, the ends (telomeres) shorten. In growing cells, the enzyme telomerase replaces these lost ends. Adult cells lack telomerase, limiting the number of times the cell can divide. However, telomerase is activated in cancer cells, allowing an unlimited number of cell divisions.
- Normal cells cease to divide and die when there is DNA damage or when cell division is abnormal. Cancer cells continue to divide, even when there is a large amount of damage to DNA or when the cells are abnormal. These progeny cancer cells contain the abnormal DNA; so, as the cancer cells continue to divide they accumulate even more damaged DNA.
5. What Causes Cancer?
The prevailing model for cancer development is that mutations in genes for tumor suppressors and oncogenes lead to cancer. However, some scientists challenge this view as too simple, arguing that it fails to explain the genetic diversity among cells within a single tumor and does not adequately explain many chromosomal aberrations typical of cancer cells. An alternate model suggests that there are “master genes” controlling cell division. A mutation in a master gene leads to abnormal replication of chromosomes, causing whole sections of chromosomes to be missing or duplicated. This leads to a change in gene dosage, so cells produce too little or too much of a specific protein. If the chromosomal aberrations affect the amount of one or more proteins controlling the cell cycle, such as growth factors or tumor suppressors, the result may be cancer. There is also strong evidence that the excessive addition of methyl groups to genes involved in the cell cycle, DNA repair, and apoptosis is characteristic of some cancers. There may be multiple mechanisms leading to the development of cancer. This further complicates the difficult task of determining what causes cancer.
6. Tumor Biology
Cancer cells behave as independent cells, growing without control to form tumors. Tumors grow in a series of steps. The first step is hyperplasia, meaning that there are too many cells resulting from uncontrolled cell division. These cells appear normal, but changes have occurred that result in some loss of control of growth. The second step is dysplasia, resulting from further growth, accompanied by abnormal changes to the cells. The third step requires additional changes, which result in cells that are even more abnormal and can now spread over a wider area of tissue. These cells begin to lose their original function; such cells are called anaplastic. At this stage, because the tumor is still contained within its original location (called in situ) and is not invasive, it is not considered malignant — it is potentially malignant. The last step occurs when the cells in the tumor metastasize, which means that they can invade surrounding tissue, including the bloodstream, and spread to other locations. This is the most serious type of tumor, but not all tumors progress to this point. Non-invasive tumors are said to be benign.
The type of tumor that forms depends on the type of cell that was initially altered. There are five types of tumors.
- Carcinomas result from altered epithelial cells, which cover the surface of our skin and internal organs. Most cancers are carcinomas.
- Sarcomas result from changes in muscle, bone, fat, or connective tissue.
- Leukemia results from malignant white blood cells.
- Lymphoma is a cancer of the lymphatic system cells that derive from bone marrow.
- Myelomas are cancers of specialized white blood cells that make antibodies.
Angiogenesis
Although tumor cells are no longer dependent on the control mechanisms that govern normal cells, they still require nutrients and oxygen in order to grow. All living tissues are amply supplied with capillary vessels, which bring nutrients and oxygen to every cell. As tumors enlarge, the cells in the center no longer receive nutrients from the normal blood vessels. To provide a blood supply for all the cells in the tumor, it must form new blood vessels to supply the cells in the center with nutrients and oxygen. In a process called angiogenesis, tumor cells make growth factors that induce the formation of new capillary blood vessels. The cells of the blood vessels that divide to make new capillary vessels are inactive in normal tissue; however, tumors make angiogenic factors, which activate these blood vessel cells to divide. Without the additional blood supplied by angiogenesis, tumors can grow no larger than about half a millimeter.
Without a blood supply, tumor cells also cannot spread, or metastasize, to new tissues. Tumor cells can cross through the walls of the capillary blood vessel at a rate of about one million cells per day. However, not all cells in a tumor are angiogenic. Both angiogenic and non-angiogenic cells in a tumor cross into blood vessels and spread; however, non-angiogenic cells give rise to dormant tumors when they grow in other locations. In contrast, the angiogenic cells quickly establish themselves in new locations by growing and producing new blood vessels, resulting in the rapid growth of the tumor.
How do tumors begin to produce angiogenic factors? An oncogene called BCL2 has been shown to greatly increase the production of a potent stimulator of angiogenesis. It appears, then, that oncogenes in tumor cells may cause an increased expression of genes that make angiogenic factors. There are at least fifteen angiogenic factors and production of many of these is increased by a variety of oncogenes. Therefore, oncogenes in some tumor cells allow those cells to produce angiogenic factors. The progeny of these tumor cells will also produce angiogenic factors, so the population of angiogenic cells will increase as the size of the tumor increases.
How important is angiogenesis in cancer? Dormant tumors are those that do not have blood vessels; they are generally less than half a millimeter in diameter. Several autopsy studies in which trauma victims were examined for such very small tumors revealed that thirty-nine percent of women aged forty to fifty have very small breast tumors, while forty-six percent of men aged sixty to seventy have very small prostate tumors. Amazingly, ninety-eight percent of people aged fifty to seventy have very small thyroid tumors. However, for those age groups in the general population, the incidence of these particular cancers is only one-tenth of a percent (thyroid) or one percent (breast or prostate cancer). The conclusion is that the incidence of dormant tumors is very high compared to the incidence of cancer. Therefore, angiogenesis is critical for the progression of dormant tumors into cancer.
7. Viruses and Cancer
Many viruses infect humans but only a few viruses are known to promote human cancer. These include both DNA viruses and retroviruses, a type of RNA virus. (See the HIV and AIDS unit.) Viruses associated with cancer include human papillomavirus (genital carcinomas), hepatitis B (liver carcinoma), Epstein-Barr virus (Burkitt’s lymphoma and nasopharyngeal carcinoma), human T-cell leukemia virus (T-cell lymphoma); and, probably, a herpes virus called KSHV (Kaposi’s sarcoma and some B cell lymphomas). The ability of retroviruses to promote cancer is associated with the presence of oncogenes in these viruses. These oncogenes are very similar to proto-oncogenes in animals. Retroviruses have acquired the proto-oncogene from infected animal cells. An example of this is the normal cellular c-SIS proto-oncogene, which makes a cell growth factor. The viral form of this gene is an oncogene called v-SIS. Cells infected with the virus that has v-SIS overproduce the growth factor, leading to high levels of cell growth and possible tumor cells.
Viruses can also contribute to cancer by inserting their DNA into a chromosome in a host cell. Insertion of the virus DNA directly into a proto-oncogene may mutate the gene into an oncogene, resulting in a tumor cell. Insertion of the virus DNA near a gene in the chromosome that regulates cell growth and division can increase transcription of that gene, also resulting in a tumor cell. Using a different mechanism, human papillomavirus makes proteins that bind to two tumor suppressors, p53 protein and RB protein, transforming these cells into tumor cells. Remember that these viruses contribute to cancer, they do not by themselves cause it. Cancer, as we have seen, requires several events.
8. Environmental Factors
Several environmental factors affect one’s probability of acquiring cancer. These factors are considered carcinogenic agents when there is a consistent correlation between exposure to an agent and the occurrence of a specific type of cancer. Some of these carcinogenic agents include X-rays, UV light, viruses, tobacco products, pollutants, and many other chemicals. X-rays and other sources of radiation, such as radon, are carcinogens because they are potent mutagens. Marie Curie, who discovered radium, paving the way for radiation therapy for cancer, died of cancer herself as a result of radiation exposure in her research. Tobacco smoke contributes to as many as half of all cancer deaths in the U.S., including cancers of the lung, esophagus, bladder, and pancreas. UV light is associated with most skin cancers, including the deadliest form, melanoma. Many industrial chemicals are carcinogenic, including benzene, other organic solvents, and arsenic. Some cancers associated with environmental factors are preventable. Simply understanding the danger of carcinogens and avoiding them can usually minimize an individual’s exposure to these agents.
The effect of environmental factors is not independent of cancer genes. Sunlight alters tumor suppressor genes in skin cells; cigarette smoke causes changes in lung cells, making them more sensitive to carcinogenic compounds in smoke. These factors probably act directly or indirectly on the genes that are already known to be involved in cancer. Individual genetic differences also affect the susceptibility of an individual to the carcinogenic affects of environmental agents. About ten percent of the population has an alteration in a gene, causing them to produce excessive amounts of an enzyme that breaks down hydrocarbons present in smoke and various air pollutants. The excess enzyme reacts with these chemicals, turning them into carcinogens. These individuals are about twenty-five times more likely to develop cancer from hydrocarbons in the air than others are.
9. Detecting and Diagnosing Cancer
The most common techniques for detecting cancer are imaging techniques such as MRI, X-rays (such as mammograms), CT, and ultrasound, which can provide an image of a tumor. Endoscopy allows a physician to insert a lighted instrument to look for tumors in organs such as the stomach, colon, and lungs. Most of these techniques are used to detect visible tumors, which must then be removed by biopsy and examined microscopically by a pathologist. The pathologist looks for abnormalities in the cells in terms of their shape, size, and structure, especially the nucleus. In addition, the pathologist looks at the borders of the tumor to see whether those cells are normal. Based on examination of the tumor cells, the pathologist determines whether the tumor is benign or malignant, and determines whether is in an early or late stage of development. Diagnosis may also include the removal and examination of lymph nodes to determine whether the cancer cells have spread.
Tumor markers are proteins found more often in the blood of individuals with the tumor than in normal individuals. These are not ideal compounds for diagnosing of cancer for two reasons. First, individuals without cancer may have elevated levels of the marker, leading to false positives. Second, tumor markers are not sufficiently elevated in all individuals with cancer to allow their detection. This leads to false negatives. One of the most commonly used tumor markers is prostate-specific antigen (PSA). It is present in all adult males, but its level is increased after both benign and malignant changes in the prostate. Therefore, high levels of PSA indicate only that further tests are required to determine whether the condition is cancer. If prostate cancer is diagnosed, the levels of PSA can help to determine the effectiveness of treatment and detect recurrence. Another tumor marker is CA 125, which is produced by a number of different cells, particularly ovarian cancer cells. It is used primarily to monitor the treatment efficacy of ovarian cancer. When the cancer is responding to treatment, CA 125 levels fall. It is not used as a routine test for ovarian cancer because many common conditions that cause inflammation also increase the level of CA 125, leading to a high incidence of false positives.
The earlier a cancer is found the more effectively it can be treated; however, early-stage cancers typically produce no symptoms. Scientists are developing molecular techniques to detect very early cancer. Using techniques such as mass spectrometry, they are also developing specific blood tests to identify a pattern of new proteins in the blood of individuals with a particular type of cancer. (See the Proteomics unit.) In addition, scientists are developing DNA microarrays to identify genes expressed in particular types of cancer cells. (See the Genomics unit.)
With the sequencing of the human genome and the mapping of single nucleotide polymorphisms (SNPs) (see the Genomics unit), it may be possible to diagnose particular cancers by identifying cells with known gene alterations. In 2002 scientists detected ovarian cancer by testing blood for the presence of DNA released by tumor cells. They looked for changes in certain alleles at eight SNPs that are characteristic of cancer. Using this technique, they successfully identified eighty-seven percent of patients known to have early-stage of ovarian cancer and ninety-five percent of those with late-stage ovarian cancer. The ability to determine which genetic alterations are associated with various cancers opens up the possibility of identifying cancerous cells while the cancer is in an early, treatable stage.
10. Traditional Treatments
Because cancer comprises many diseases, doctors use many different treatments. The course of treatment depends on the type of cancer, its location, and its state of advancement. Surgery, often the first treatment, is used to remove solid tumors. It may be the only treatment necessary for early stage cancers and benign tumors. Radiation kills cancer cells with high-energy rays targeted directly to the tumor. It acts primarily by damaging DNA and preventing its replication; therefore, it preferentially kills cancer cells, which rapidly divide. It also kills some normal cells, particularly those that are dividing. Surgery and radiation treatment are often used together.
Chemotherapy drugs are toxic compounds that target rapidly growing cells. Many of these drugs are designed to interfere with the synthesis of precursor molecules needed for DNA replication; they interfere with the ability of the cell to complete the S phase of the cell cycle. Other drugs cause extensive DNA damage, which stops replication. A class of drugs called spindle inhibitors stops cell replication early in mitosis. During mitosis, chromosome separation requires spindle fibers made of microtubules; spindle inhibitors stop the synthesis of microtubules. Because most adult cells don’t divide often, they are less sensitive to these drugs than are cancer cells. Chemotherapy drugs also kill certain adult cells that divide more rapidly, such as those that line the gastrointestinal tract, bone marrow cells, and hair follicles. This causes some of the side effects of chemotherapy, including gastrointestinal distress, low white blood cell count, and hair loss.
11. Newer Treatments
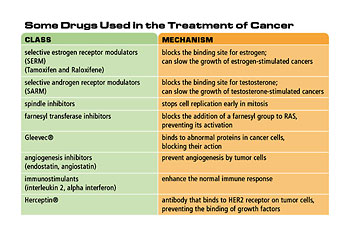
Many of the factors that affect normal cell growth are hormones. Although cancer cells have lost some of the normal responses to growth factors, some cancer cells still require hormones for growth. Hormone therapy for cancer attempts to starve the cancer cells of these hormones. This is usually done with drugs that block the activity of the hormone, although some drugs can block synthesis of the hormone. For example, some breast cancer cells require estrogen for growth. Drugs that block the binding site for estrogen can slow the growth of these cancers. These drugs are called selective estrogen receptor modulators (SERMs) or anti-estrogens. Tamoxifen and Raloxifene are examples of this type of drug. A ten-year clinical trial of these two drugs with 20,000 women began in 1999 to determine their effectiveness in preventing breast cancer. Similarly, testosterone (an androgen hormone) stimulates some prostate cancer cells. Selective androgen receptor modulators (SARMs) are drugs that block the binding of testosterone to these cancer cells, inhibiting their growth and possibly preventing prostate cancer.
Newer chemotherapeutic drugs target specific, active proteins or processes in cancer cell signal transduction pathways, such as receptors, growth factors, or kinases (see Fig. 1). Because the targets are cancer-specific proteins, the hope is that these drugs will be much less toxic to normal cells than conventional cancer drugs.
The oncogene RAS is mutated in many types of cancer, particularly pancreatic cancer, which has a poor rate of survival for those afflicted. The RAS protein is only active after it is modified by the addition of a specific chemical group. Scientists are developing drugs to inhibit the action of the enzyme that adds the chemical group to the RAS protein, resulting in an inactive form of RAS. Early tests indicate that these drugs show promise for reducing tumors in cancer patients.
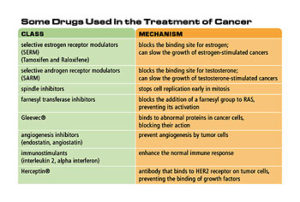
Table 2. Some drugs used in the treatment of cancer
A drug called Gleevec® inhibits cancer cell growth and causes cancer cells to undergo apoptosis, or programmed cell death. It binds to abnormal proteins in cancer cells, blocking their action in promoting uncontrolled cell growth. Because it binds only to these abnormal proteins, Gleevec® does not show the high levels of toxicity of other chemotherapy drugs. Gleevec® was developed to treat a relatively rare cancer called chronic myeloid leukemia; however, it also appears to help other cancers.
Chemotherapy may fail because the cancer cells become resistant to the therapeutic drugs. One of the characteristics of cancer cells is a high frequency of mutation. In the presence of toxic drugs, cancer cells that mutate and become resistant to the drug will survive and multiply in the presence of the drug, producing a tumor that is also resistant to the drug. To overcome this problem, combinations of chemotherapy drugs are given at the same time. This decreases the probability that a cell will develop resistance to several drugs at once; however, such multiple resistances do occur. Some drug-resistant cancer cells express a gene called MDR1 (multiple drug resistance). This gene encodes a membrane protein that can not only prevent some drugs from entering the cell, but can also expel drugs already in the cell. Some cancer cells make large amounts of this protein, allowing them to keep chemotherapy drugs outside the cell.
Another promising target for cancer therapy is angiogenesis. Several drugs, including some naturally occurring compounds, have the ability to inhibit angiogenesis. Two compounds in this class are angiostatin and endostatin; both are derived from naturally occurring proteins. These drugs prevent angiogenesis by tumor cells, restricting tumor growth and preventing metastasis. One important advantage of angiogenesis inhibitors is that, because they do not target the cancer cells directly, there is less chance that the cancer cells will develop resistance to the drug.
One contributing factor in cancer is the failure of the immune system to destroy cancer cells. Immunotherapy encompasses several techniques that use the immune system to attack cancer cells or treat the side effects of some types of cancer treatment. The least specific of these are the immunostimulants, such as interleukin 2 and alpha interferon, which enhance the normal immune response.
A technique called chemoimmunotherapy attaches chemotherapy drugs to antibodies that are specific for cancer cells. The antibody then delivers the drug directly to cancer cells without harming normal cells, reducing the toxic side effects of chemotherapy. These molecules contain two parts: the cancer-cell-specific antibody and a drug that is toxic once it is taken into the cancer cell. A similar strategy radioimmunotherapy, couples specific antibodies to radioactive atoms, thereby targeting the deadly radiation specifically to cancer cells.
Another immunological approach uses antibodies that inactivate cancer-specific proteins, such as growth factors or tumor cell receptors, which are required by tumor cells. For example, many breast and ovarian cancer cells over-express a receptor protein called HER2. An antibody called Herceptin, which binds HER2, inhibits tumor growth by preventing the binding of growth factors to these cells.
Some cancers, particularly leukemia, are treated with very high doses of chemotherapy drugs and radiation intended to kill all the cancer cells. The side effect of this harsh treatment is the destruction of the bone marrow, which contains stem cells. Stem cells, immature cells that develop into blood cells, are essential. After treatment, the patient’s bone marrow must be restored, either from bone marrow removed from the patient before drug therapy or from a compatible donor. Although the patient’s own bone marrow is best, it can contain cancer cells that must be destroyed before it is returned to the patient.
12. Preventing Cancer
Cancer appears to result from a combination of genetic changes and environmental factors. A change in lifestyle that minimizes exposure to environmental carcinogens is one effective means of preventing cancer. Individuals who restrict their exposure to tobacco products, sunlight, and pollution can greatly decrease their risk of developing cancer. Many foods contain antioxidants and other nutrients that may help to prevent cancer. The National Cancer Institute recommends a diet with large amounts of colorful fruits and vegetables. These foods supply ample amounts of vitamin A, C, and E, as well as phytochemicals and other antioxidants that help to prevent cancer. There is strong evidence that a diet rich in vegetables and fruits will not only reduce the risk of cardiovascular disease, obesity, and diabetes, but will also protect against cancer.
Vaccines also offer some promise for the prevention of cancer. The first vaccine to prevent cancer was for hepatitis B, which is associated with liver cancer. An effective hepatitis B vaccine is available that can prevent both hepatitis and the cancer that may follow this infection. In 2002, test results of a papillomavirus vaccine were reported. Human papillomavirus type 16 infects about twenty percent of adults. Although most papillomavirus infections do not cause cancer, some are associated with cervical cancer. A vaccine against this virus was administered to 1,200 young women in the United States. Within eighteen months, the vaccine produced high levels of antibodies to the virus, and prevented both papillomavirus infection and precancerous lesions in all the women. In the control group of about
1,200 women who did not receive the vaccine, forty-one infections and nine precancerous lesions were found. The vaccine can also prevent genital warts caused by this virus strain. It appears that vaccines such as these may help in the fight to prevent cancers associated with viruses.
13. Screening, Genetic Tests, and Counseling
Early diagnosis of cancer greatly increases survival; therefore, regular exams for cancer can help to prevent deaths from cancer. These include mammograms and Pap tests for women, prostate cancer tests for men, colonoscopy exams for colon cancer, and regular physical exams for other types of cancer. Individuals with a strong family history of cancer should consider genetic tests for cancer and cancer risk counseling. The focus of cancer risk counseling is the individual’s personal risk of developing cancer and appropriate actions based on that risk.
The discovery of the BRCA1 and BRCA2 genes associated with early development of breast cancer has allowed women with a family history of early breast cancer to be tested for mutations in these genes. Only five to ten percent of breast cancers show evidence of inheritance. Of these, forty-five percent are associated with a mutation in BRCA1 and thirty-five percent with BRCA2. The gene or genes for the remaining twenty percent are not yet known. If the BRCA1 and BRCA2 test results are negative, there is no evidence that the woman will have breast cancer because of these mutations. However, she may get breast cancer because of somatic mutations in these or other genes. If the BRCA1 or BRCA2 test is positive, other family members may be tested to determine whether the gene was inherited. If other family members are negative, then there is less chance of hereditary risk of this form of cancer, although the individual with the mutation does carry an increased risk of the disease. If the test is positive in other family members, there is an increased hereditary risk for breast cancer in that family. The absence of hereditary risk does not mean that there is no other risk for breast cancer.
Decisions based on genetic tests can be very complicated. Individuals must be fully informed about the risks before they can make reasonable decisions. Genetic counselors are trained to help individuals make difficult decisions based on genetic tests. The cumulative risk of breast cancer to age seventy for a woman with a BRCA1 mutation is about fifty-seven to eighty-five percent depending on whether she is in a high-risk family. Some women find the fear of cancer so disruptive to their lives that they choose mastectomy to prevent cancer. (This is called prophylactic mastectomy.) Similarly, women with BRCA1 have a high lifetime risk of ovarian cancer, causing some of them to choose to have their ovaries removed. While these are difficult decisions, the availability of genetic information provides individuals with information that they can use to make such important medical decisions. A young woman with a strong family history of ovarian cancer might find by genetic testing that she does not have the BRCA1 mutation and should not consider removal of her ovaries.