




Rediscovering Biology: Molecular to Global Perspectives
Online Textbook and Video
This online textbook chapter supports and extends the content of the Sex and Gender video. The chapter covers genetic imprinting, inter-sex conditions, the effect of hormones on sex determination, and ethics. Click on the Go button to begin reading, or skip to a sub-section of the chapter in the list below.
1. Introduction
“I think humans like things to be ordered, and they get bothered about gray areas and when things become less clear-cut. But these days I don’t think so much in black and white about male and female. Now I think of it all as being on a spectrum.”
– Dr. Andrew Sinclair 1
Introduction:
Max Beck was born an intersexual, someone with ambiguous genitals. Like most babies without a normal penis, he was “assigned” the sex of female and underwent plastic surgery to “fix” his genitals. He was named Judy and grew up as a girl, a self-described tomboy. In his teens, more surgery and female hormone injections turned him into a woman – a woman with no sense of gender identity. As a young adult he had sexual relationships with males and females, first accepting himself as a lesbian, then marrying a man. After divorcing his husband, he once again became a lesbian with a partner. Finally he received his medical records, which revealed that he was an intersexual and had both an X and a Y chromosome. Over a period of two years he decided that he could no longer live as a female. He reassigned himself as a male, married his female partner, and became the father of a child, conceived by his wife using donor sperm. Despite his sex assignment as a female at birth, Max was never able to accept his gender as female.
In contrast, Jan Morris was born James Morris, an apparently normal male. A successful journalist, author, and mountain climber, she married and had five children before she decided in her 30s to change her sex to female and her name to Jan. Jan Morris wrote of her sex change in the book, Conundrum, explaining that she had always known that she was a woman, wrongly born into the body of a man. She has continued to be a successful writer and lives harmoniously with her former wife.
The use of pronouns above may strike some as strange. Which pronoun should be used in the case of transgendered individuals? This question highlights the difficulties our language and culture have in confronting issues of sex and gender. We have used the pronoun of the individual’s final choice of gender.
What is the difference between sex and gender? Max Beck was born a male (albeit with ambiguous genitals), and efforts to change him into a girl by surgery and hormones failed to change his gender, his sense of identity. In contrast, Jan Morris believes that she was born a woman in a man’s body. Transsexuals like Jan Morris explain that they must change their physical sex because there is no way to change their gender, which comes from the brain. While there is no general agreement on terminology, the Merriam-Webster dictionary defines gender as “the behavioral, cultural, or psychological traits typically associated with one sex,” and defines sex as “either of the two major forms of individuals that occur in many species and that are distinguished respectively as female or male.” This suggests that sex is biological while gender is subjective.
2. Sex and the Y Chromosome
Except for the sex chromosomes (X and Y), all humans have the same set of chromosomes. The karyotype of a human male is 46XY (46 chromosomes, including one X and one Y), and that of a female is 46XX (46 chromosomes, including two X chromosomes.) In mammals as a whole, the presence or absence of the Y chromosome usually determines sex. Individuals with an X chromosome but no Y (45XO) are female (Turner’s syndrome); individuals with two X chromosomes and a Y chromosome (47XXY) are male (Kleinfelter’s syndrome).
The Y chromosome is considerably smaller than the X chromosome and has a much lower density of genes. In fact, the Y has often been called a “genetic junkyard.” But there are a few rubies among the rubbish of that genetic junkyard: the Y chromosome contains the genes are essential for male fertility and other male characteristics.
Why does the Y chromosome have so few functional genes? Evolutionary biologists are still debating the details but they agree that the lack of recombination explains the paucity of functional genes on the Y. Unlike the twenty-two pairs of autosomes, there is no recombination between the X and most of the Y chromosome. Genes on the part of the Y chromosome that does not recombine will be passed from father to son, down a paternal lineage, and will never be present in females. The lack of recombination weakens the effectiveness of natural selection to weed out bad variants and select for good ones. Over many millions of years mutations and random genetic drift erode the Y chromosome, turning it into a genetic junkyard. In contrast, genes on the X are present in both males and females; X chromosomes, like autosomes, recombine in production of female gametes.

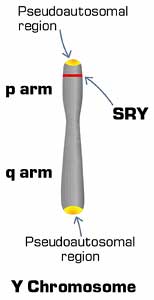
Figure 1. The Y chromosome is very small compared to the X chromosome. The pseudoautosomal regions at the tips contain the genetic material on the Y that shows similarity to the X chromosome. The SRY gene is located on the p arm of the Y.
About five percent of the Y chromosome does recombine with the X. This region, at the tips of the chromosomes, is called the pseudoautosomal region because in it the X and Y chromosomes behave as autosomes (Fig. 1). The pseudoautosomal region is more gene-rich than the rest of the Y chromosome. Several of the genes on the pseudoautosomal region of the Y have counterparts on X, reflecting a common evolutionary ancestor. The genes required for male fertility are found in the non-recombining regions of the Y, and are not present on X.
Researchers in David Page’s lab have shown that one-quarter of the Y chromosome consists of eight families of nearly identical nucleotide sequences, and includes duplicate copies of important genes. Because these regions are arranged in palindrome fashion, they provide a mechanism for a kind of internal recombination between the similar genes on the same chromosome. This process, called gene conversion, aids in the detection and repair of gene mutations in this part of the Y chromosome.2
3. Paternal Inheritance

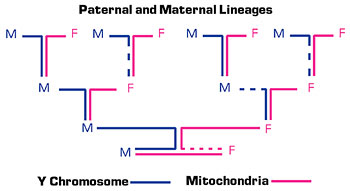
Figure 2. Maternal lineages can be traced through mitochondrial genes, which are inherited by males and females only from the mother. Paternal lineages can be traced through the Y chromosome, which is inherited only by males and only from the father. (M=male and F=female)
The lack of recombination means that the entire non-recombining portion of the Y is passed intact from father to son. A male shares the same Y chromosome with his father, paternal grandfather, paternal great-grandfather, and so on. (Fig. 2). Researchers can establish paternal genetic relationships by comparing small differences (polymorphisms) between modern Y chromosomes. The identification of genetic markers such as single nucleotide polymorphisms">single nucleotide polymorphisms (SNPs) and indels (insertions and deletions) in the non-recombining regions of the Y provides a tool to study population structure and history, genealogy, and human evolution. Because these regions do not recombine they change very slowly, so they may be useful in identifying stable paternal lineages over thousands of years. Mutations occasionally occur in this DNA, however, which are then inherited down the paternal line.
4. Evolution of the Y Chromosome
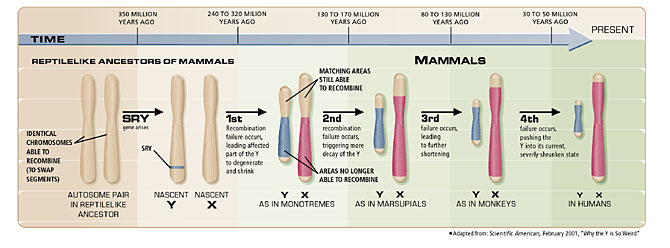
The evolutionary ancestor of the sex chromosomes was a pair of matched, autosomal chromosomes that acquired sex-determining genes on one member of the pair. This occurred about 350 million years ago in a reptile-like ancestor. Over time additional genes with male-specific functions accumulated in this same chromosome, called proto-Y, which then lost its ability to recombine with its counterpart chromosome, called proto-X. There are four regions of the proto-X chromosome, which appear to have been involved in four different steps, resulting in the loss of recombination with proto-Y. Each of the four regions accumulated mutations in those non-recombining regions of proto-Y at four different times in evolution. Each time recombination was lost there was degradation and loss of the non-recombining region. Over time this chromosome evolved into Y, losing most of its genetic information as a result of the degradation of the non-recombining regions of the chromosome. Its partner chromosome evolved into the X chromosome. The degeneration of the Y was offset at various times by additions of autosomal genes to this chromosome (as well as to X), leading to a pattern of loss and gain of genetic material over a period of about 170 million years (Fig. 3).

Figure 3. The degeneration of the Y occurred in four discrete episodes, beginning about 300 million years ago when a reptile-like ancestor acquired the SRY gene on one of its autosomal chromosomes. Each of the four episodes involved a failure of recombination to occur between the X and the Y chromosomes, resulting in subsequent decay of some genes in the non-recombining region.
5. X Inactivation
Having a single copy of any chromosome other than the X or the Y is lethal in humans; however, only one X chromosome is needed for normal development to occur. Therefore, the evolutionary process that resulted in a loss of genes from the Y chromosome would seem to have presented a problem. At least two possible mechanisms could balance gene expression between the two X chromosomes in females, versus only one X in males. Gene activity on the one X present in males (relative to the ancestor before the evolution of the XY system) could be increased so that these genes produce twice as much in males as in females. Alternately, X-linked genes could have their activity decreased in females. The first mechanism is seen in some insects, including Drosophila, while mammals use a special variant of the second, called X-inactivation. In X-inactivation, female embryos randomly inactivate one X chromosome in each cell, resulting in only one functional copy of X-linked genes in both males and females.
X-inactivation requires a locus on the X, called the X-inactivation center. At this locus, inactivation occurs in response to a developmental cue, which is present only at specific stages of embryo development. Inactivation occurs because of a specific type of RNA, which binds to one X chromosome, preventing transcription of the genes on this particular copy. In addition, enzymes add methyl groups to the DNA of the inactive X, resulting in repression of transcription. The inactivated X is visible during interphase in mitosis as a condensed chromosome, called a Barr body. It replicates in the S (synthesis) phase of the cell cycle later than does the active copy. Inactivation of one of the two X copies in a female leaves only one active
X chromosome in any cell. An individual who has three X chromosomes has two inactivated copies of the X, producing two Barr bodies.
Because the X is inactivated randomly in cells, one cell could have the maternal X inactivated, while the adjacent cell could have the paternal X inactivated. This causes a pattern of gene expression called mosaicism">mosaicism, which occurs when different alleles of X-linked genes are expressed in different cells. A classic example of mosaicism is the female calico cat, which inherits an X-linked allele for yellow coat color from one parent and an X-linked black allele from the other. One or the other color is expressed in patches of the coat that represent cells descending from parental cells with either an active maternal X or paternal X.
6. Genetic Imprinting
Some genes are expressed only from the maternal chromosome, while others genes are expressed only from the paternal chromosome. The second gene copy is silenced during gamete formation in the egg (when maternal gene copies are silenced) or the sperm (when paternal gene copies are silenced). This is known as genetic imprinting. Imprinting occurs in each generation when new egg and sperm cells are produced.
Relatively few genes in humans are known to be imprinted and such genes tend to be clustered in the genome. The gene imprinting occurs by the addition of methyl groups to the DNA of the silenced gene, preventing transcription of the gene. This gene silencing acts in much the same manner as mutation or deletion of one copy of a gene, except that it is not a permanent, heritable change. If the one remaining active gene is deleted or mutated, there is no extra, functional copy on the second chromosome; therefore, mutation of the single, active copy of an imprinted gene may result in disease. Similarly, as a result of an error, cells may receive all or part of a pair of chromosomes from a single parent. For imprinted genes, that means that the cell receives either two imprinted copies or two active copies. If both copies are imprinted, there is no functional gene. Two active copies of a gene may also result from a mutation that leads to loss of imprinting; neither copy will be silenced. Too many active copies of a gene may result in overexpression of a gene, which can cause disease. A number of cancers have been associated with failure to imprint genes, especially genes that produce growth factors. Overexpression of growth factors can disrupt the cell cycle, contributing to uncontrolled cell growth and cancer. (See the Cell Biology and Cancer unit.)
7. Testis-Determining Factor
The presence of a Y chromosome is usually necessary and sufficient for male development: a 45XO human is female, while a 47XXY is male. It also typically leads to formation of a testis in the mammalian embryo – the primary sex-determining event. The testis then produces and secretes the male hormones, androgens, resulting in the formation of male genitalia. In the absence of Y, the pathway leads to development of a female (Fig. 4). Therefore, the Y must contain a testis-determining factor.

Figure 4. The genital ridge in an embryo is converted to a bipotential gonad by the products of the LHX9, SF1, and WT1 genes. This gonad develops into an ovary under the influence of the WNT4 and DAX1 gene products; it develops into a testis under the influence of the products of the SRY and SOX9 genes. The ovary produces cells that make estrogen, which causes the Müllerian duct to differentiate into the female genitalia. The testis makes two hormones, anti-Müllerian duct factor (AMH), which causes the Müllerian duct to regress, and testosterone, which causes the Wolffian duct to differentiate into male internal organs. Testosterone is also converted into dihydrotestosterone (DHT), which is required for development of male external genitalia.

Table 1. A chart of the genes that are involved in human sex determination.
The region of the Y chromosome that carries the testis-determining factor contains a gene called SRY (sex region Y). Its product binds to DNA, acting as a transcription factor that is critical for testis production. Scientists studying sex reversal, a difference between the chromosomal sex and the phenotypic sex, confirmed the importance of SRY. They determined that infertile males who were XX had all acquired a particular snippet of the Y chromosome, which was translocated to X. That small fragment of the Y carries SRY. Conversely, many XY females have a deletion of the part of the Y that includes SRY. Introduction of the mouse SRY gene into an XX mouse causes the formation of testis and the animal develops as a male anatomically; however, it does not produce sperm. Thus, SRY is the testis-determining factor, and is the only gene on the Y chromosome that is essential for development of male genitalia. Some genes required for male fertility are on the Y chromosome, while others are on the X or on autosomal chromosomes. The DAZ genes on the Y are essential for sperm formation; deletion of DAZ results in male infertility.
In the first few weeks of development a human embryo develops a sexually indifferent gonad, which can become either a testis or an ovary. Without SRY to stimulate testis development, the gonad becomes an ovary and the embryo develops into a female; the development pathways of both male and female are complex, however, and are regulated by several gene products (Fig. 4). For example, the product of the DAX1 gene (present on the X chromosome) appears to interact with SRY: an excess of SRY leads to testis formation, while an excess of DAX1 leads to ovary formation. A mutation in DAX1 leads to sterile males but has no effect on females. An extra copy of DAX1 in a male leads to a sex-reversed XY female. An SOX9 mutation (on chromosome 17) in a male leads to sex-reversed XY females, while an extra copy of SOX9 in a female can result in a sex-reversed XX male. Conversely, an extra copy of WNT4, which is implicated in ovary formation, in a male results in a sex-reversed XY female.
8. Hormones
Hormones are small molecules that bind to specific target cells to modify the response of the cell, usually affecting gene expression. For example, estrogen is a small, hydrophobic molecule that binds to estrogen receptors. The estrogen-receptor complex then enters the nucleus and binds to specific DNA sequences in certain genes, and turns on or off transcription of those genes.

Table 1. Genes Involved in Human Sex Determination
A chart of the genes that are involved in human sex determination.
In females the ovaries secrete estrogens and progesterone, which are essential for the development of female genitalia during fetal development (Fig. 4). These hormones are also required for sexual development at puberty, and for pregnancy. The ovaries also produce a small amount of testosterone, although much less than testes in males produce.
In males the testes secrete the major androgen, testosterone. Synthesis of this hormone increases significantly at puberty, when it is responsible for adult sexual development. Androgens are also essential for the development of male genitalia during fetal development (Fig. 4). Some testosterone is converted to estrogen in males and is important for bone formation.
Animals in utero can be affected by hormones produced by nearby siblings of the opposite sex. The placement of an animal, such as a mouse in a litter, may have a long-term effect on physiology or behavior. Female mice that develop in the uterus between two males have shorter fertile periods than do females that develop between two females. Male mice prefer to mate with the females that develop in an all-female environment. Females who develop between two brothers in utero are more aggressive towards intruders than are females who develop with two sisters.
Hormones also affect mature adults. Males and females have receptors for estrogens, progesterone, and androgens in various tissues. Transsexuals (individuals who have a conflict between their biological sex and their perceived gender) must take hormones of the opposite sex if they choose to undergo a sex change. Males can develop breasts, decrease facial hair production, and change the texture of their skin and hair as a result of estrogen and progesterone therapy combined with anti-androgen drugs. Conversely, high levels of testosterone can have a masculizing effect on females. Interestingly, individual differences in natural hormone levels and hormone sensitivity mean that those undergoing a sex change require individualized hormone treatment programs.
9. Intersex
For some individuals, determination of biological sex can be difficult. Intersex refers to genetically determined differences of the reproductive system. This can include differences in internal reproductive organs, external genitalia, or karyotype. Mild intersex conditions include, in males, a condition in which the urethra opens on the underside of the penis or, in females, an enlarged clitoris. Female intersexuals (karyotype 46XX) (also called female pseudohermaphrodites) have normal ovarian tissue, and have either male or ambiguous genitalia. This is usually a result of a change in the fetal adrenal glands, leading to production of abnormally high levels of androgens. The androgens produce some masculine features in female infants: ovaries and uterus form, but the external genitalia appear male-like. This accounts for about two-thirds of intersex states.
Male intersexuals (karyotype 46XY) (also called male pseudohermaphrodites) have normal testes with female or ambiguous genitalia. They most often result from several different genetic alterations in pathways of testosterone synthesis and metabolism. For example, males who have a mutation in the gene that converts testosterone to dihydroxytestosterone have normal testes but have a very small penis and a vaginal pouch. In gonadal dysgenesis the testes fail to secrete androgens or Müllerian-inhibiting hormone, leading to formation of female genitalia. With estrogen treatment, however, these individuals will grow into females. A condition called micropenis results from lack of androgens later in fetal life; testosterone treatment can stimulate masculizing puberty in these individuals.
Androgen insensitivity syndrome (AIS) occurs when a male produces cells that cannot respond to androgen. The defect is in a gene on the X chromosome that produces the androgen receptor. Individuals may have complete or partial androgen insensitivity. In complete AIS the testes develop in the embryo, and produce testosterone and the hormone that inhibits development of female internal reproductive organs (Fig. 4). However, because the cells do not respond to testosterone, female genitals develop, which may be incomplete. The newborn appears to be a female and develops external female characteristics at puberty. Lacking internal female reproductive organs, though, the individual with AIS does not menstruate and is infertile. In incomplete AIS, individuals may appear male or female, but there may be abnormalities in the external genitalia. Maria Patino, a Spanish runner with complete AIS, was not allowed to compete in the 1985 World University Games in Kobe, Japan because she failed the gender test. (See the Sex and Gender video.) Because of such difficulties in determining sex, the International Olympic Committee abolished gender testing in 1999.
10. Ethics of Intersex Treatment
Common medical treatments of intersex babies include:
- Assignment of gender based on a variety of clinical tests
- Surgery to remove internal gonads that might become cancerous
- Reconstruction of external genitalia appropriate for the assigned sex
- If necessary, treatment with appropriate hormones
Unless a penis is present, most intersex babies are assigned female because it is not possible to construct a fully functional penis. The gender assignment and the surgery are usually done immediately after birth, with possible additional surgery after puberty. Today, some physicians such as Eric Vilain (featured in the video) recommend allowing the child to make his or her own surgical decisions later in life. However, most parents choose the surgery earlier because they are uncomfortable with the ambiguity.
The Intersex Society of North America believes that intersex is not an abnormality but rather “an anatomical variation from the standard male and female types.”3 The Society also believes that the decision regarding treatment, if any, should be made by the individual when he or she is capable of informed consent. The Society has two objections to treatment:
1) treatment assumes that intersexuality is a disease, and 2) surgery often damages sexual function, while still failing to produce anatomically normal genitals. They claim that physicians have traditionally failed to communicate to parents the basis for the assignment of gender, which is not always (and in the case of XY individuals without a penis, never) made based on biological sex. They also claim that some physicians have failed to inform parents of alternatives and presented elective surgery as essential for health.
Physicians were once guided by the idea that infants are gender neutral at birth, and that normal gender development would follow from the environment of the child based on the sex assigned to the child.4 They believed that gender came from “nurture” rather than “nature.” In recent years individuals, especially intersexuals, have protested that the environment does not control gender and, given that the treatments are irreversible, they should be delayed until the child determines his or her true gender. Others worry, however, that children with ambiguous genitals will be ridiculed, causing permanent psychological damage that could be avoided.
11. Homosexuality
There is no simple genetic test to differentiate homosexuals from heterosexuals. However, many studies of siblings have consistently found a much higher incidence of homosexuality in pairs of monozygotic (identical) twins compared to dizygotic (fraternal) twins, strongly suggesting a genetic component to homosexuality. Although there is some suggestive evidence linking variation at specific regions of the genome with the propensity for being homosexual, these studies are not yet conclusive.
12. Sex and Disease
Sex is an important aspect of human identity, but it is also important in health. Women outlive men. In the United States at the start of the twenty-first century, a woman’s life expectancy at birth is 79 years, and a man’s is 72. While other countries have greater or lesser average life expectancies, female life expectancy is still greater in nearly all countries. In fact, for most animalsthat have been studied, females outlive males; female sperm whales outlive males by thirty years on average. Many factors contribute to this effect, including genes, hormones, and lifestyle factors. Historically, the greatest death risk for women has been childbirth. In developed countries, however, this risk has decreased markedly in the last century, significantly increasing a woman’s lifespan.
Males die more often than females – even before birth. Although there are 115 male fertilized eggs for every 100 female, the ratio for live births is
104 males to 100 females. Each year after birth, more males die than females; so, by age 100 there are only 11 males for every 100 females. With improvements in health care, the gap between longevity in men and women is decreasing. However, one troubling factor contributing to the narrowing of the gap is an increase of diseases in women that have been typically considered male diseases, especially cardiovascular disease.
There are two aspects of the longevity gap: Why do men die young and why do women live so long? Hormones appear to be part of the answer to both of these questions. Testosterone may contribute to early death in males. The greatest difference in death rates between males and females occurs during the teen years, when males experience a surge in testosterone. This increase correlates to increases in death in males by accidents, homicide, and suicide; however, these behavior-related deaths continue to contribute throughout life to male mortality more than they do to female mortality.
While teenage females also die from behavioral causes, the incidence is much lower than for males. Female teenagers also experience an increase in hormones; these hormones, however, generally correlate with increased longevity in women. The strongest evidence for the protective effects of female hormones is the increased risk for several diseases after menopause, notably cardiovascular disease and osteoporosis. In males of all ages, testosterone increases the levels of undesirable LDL cholesterol and decreases the levels of the desirable HDL cholesterol, increasing the risk for cardiovascular disease. In contrast, estrogen appears to have a beneficial effect on cholesterol levels. As of 2003, there is much controversy about whether estrogen replacement after menopause gives any significant health benefits for women. In fact, some studies suggest that replacement therapy may do more harm than good.
Women may also enjoy advantages over men in physiology and metabolism, probably because of hormone differences. Women have lower metabolic rates than men, likely leading to less oxidative damage to cells. Oxidative damage results from free radicals, which alter DNA, RNA, and protein in cells. This may explain why oxidative damage is linked to diseases such as cancer, Alzheimer’s, and atherosclerosis. In animal studies, lowering metabolism by decreasing calorie consumption has been shown to significantly increase lifespan. In addition, because they menstruate, women have less iron in their blood. (High levels of blood iron are associated with oxidation of LDL cholesterol, which contributes to cardiovascular disease.)
Women also enjoy a genetic advantage because they have two copies of the X chromosome. Mutations in genes on the X chromosome typically do not cause disease in females because there is a normal copy. Two X-linked diseases are hemophilia and muscular dystrophy. Because X-inactivation occurs randomly in each cell, about half of the cells of women heterozygous for these conditions would be normal. Additionally, the normally inactivate copy of the X chromosome in females (resulting from X-inactivation during development) may be at least partially restored as women age, allowing the inactive X to provide a good copy of a gene that was lost or altered by mutation in the other X chromosome.
One area in which women do not enjoy an advantage over men is in autoimmune diseases. Women are more susceptible to these diseases, such as systemic lupus erythematosus (lupus) and rheumatoid arthritis. There isn’t a simple explanation for this increased risk; instead, it appears to result from a combination of genetic, environmental, and hormonal effects.
Lifestyle choices also affect longevity. Early in the twentieth century, men smoked more than women, a factor that is thought to account for much of the gender gap in longevity. As more women began to smoke, the gap decreased. Studies indicate that women smokers may have an increased risk of lung cancer because they have higher levels of an enzyme that produces carcinogens from tobacco smoke. In addition, middle-aged women smokers live no longer than do men smokers, suggesting that smoking eliminates any health advantage conferred by gender.
Despite the evidence for gender-based differences in physiology, metabolism, disease, and response to certain drugs, women were excluded from most medical studies for many years. Why? It wasn’t just sexism: the difficulty in controlling the monthly cycles of hormones, and the concerns about possible pregnancy simply made it easier to leave women out of the studies. In 2001 the Institute of Medicine issued the report “Exploring the Biological Contributions to Human Health: Does Sex Matter?” The report concluded that sex was very important in health, and that women should be included in all studies of diseases that could affect them.
With improvements in health care and an understanding of the importance of nutrition and exercise, it is likely that the longevity gap will continue to decrease. We may eventually understand which components of female longevity are the result of sex and which are the result of gender.
End Notes
- Sinclair, A. 2001. The Scientist 15[15]:18.
- Rozen, S., H. Skaletsky, J. D. Marszalek, P. J. Minx, H. S. Cordum, R. H. Waterson, R. K. Wilson, and D. C. Page. 2003. Abundant gene conversion between arms of palindromes in human and ape Y chromosomes. Nature 423:873-76.
- http://www.isna.org/library/dreger-compare.html
- Money, J., J.G. Hampson, and J.L. Hampson, 1955. Hermaphroditism: Recommendations concerning assignment of sex, change of sex and psychological management. Bulletin of the Johns Hopkins Hospital 97:284.