



Rediscovering Biology: Molecular to Global Perspectives
Online Textbook and Video
This online textbook chapter supports and extends the content of the HIV and AIDS video. The chapter provides an overview of the human immune system, and covers the viral life cycle and current obstacles in controlling HIV. Click on the Go button to begin reading, or skip to a sub-section of the chapter in the list below.
1. Introduction
“The human immunodeficiency virus (HIV) epidemic has spawned a scientific effort unprecedented in the history of infectious disease research. This effort has merged aspects of clinical research, basic molecular biology, immunology, cell biology, epidemiology, and mathematical modeling in ways that have not been seen before. The ever-unfolding discoveries of novel aspects of HIV-host interaction have been accompanied by (and often have resulted from) novel interactions among researchers in the disparate disciplines.”
– John Coffin (p. 3)
In the late 1970s young homosexual men were dying from rare cancers and pneumonias caused by usually benign microbes. Such conditions, which result from failures of the immune system, became indicators of what is now called acquired immunodeficiency syndrome (AIDS). Although the causative virus, human immunodeficiency virus (HIV), was identified in 1983, there is still no cure for AIDS. In the years since, HIV has killed millions of men, women, and children from all economic classes, representing every race, from countries around the world. Each day in 2003, 15,000 more individuals became infected and 8,000 died.
HIV remains a major problem for several reasons. The virus has an extraordinarily high mutation rate, such that an infected individual often harbors many variations. This high mutation rate allows HIV to easily evolve resistance to the drugs used to treat it. In addition, cells essential to a strong immune response harbor a virus that can lay latent for years. Thus, the development of treatments and vaccines depends not only on knowledge of the complex life cycle of the virus, but also on understanding the intricate choreography of the immune system. Controlling HIV will require more than the development of medicines and vaccines, however, because poverty and politics exclude millions from treatment.
2. The Immune System
Understanding the various components of the immune system and the complex signaling that takes place between immune cells is key to understanding HIV. Both non-specific and specific lines of defense help thwart the invasion of pathogens. Non-specific defenses act quickly and indiscriminately to exclude microbes from the body or actively kill intruders. Mechanical barriers — such as the mucus, hairs, and cilia in the respiratory tract, and the flow of urine through the urinary tract — are among these non-specific defenses. Skin oils and chemicals in perspiration and gastric juices also serve as non-specific barriers. Mechanisms involving complex chemical signals such as fever and inflammation also act against a wide variety of pathogens. One non-specific defense involves phagocytes, a particular type of leukocyte (white blood cell), which act as cellular “Pac-Men,” engulfing and digesting microbes or other irritants like dust and pollen.

Table 1: Types of Leukocytes
If invaders have breached the non-specific defenses, the immune system will use a variety of leukocytes to mount directed defenses against specific invaders. Lymphocytes bind and respond to specific foreign molecules (antigens). One subset of lymphocytes, the B cells, matures into antibody-secreting cells. Another subset of lymphocytes, the T cells, includes immune cells that directly kill cancerous or virally infected cells. Some subtypes of T cells serve a regulatory function, releasing chemical signals that can stimulate or suppress a variety of immune functions. Because HIV preferentially infects one of these regulatory T cells, the so-called helper T (TH) cell, it can subvert and decimate the immune system, leading to AIDS.
3. The Central Role of Helper T Cells
Helper T (TH) cells are critical to coordinating the activity of the immune response. The chemical messages they secrete (cytokines) stimulate the non-specific immune response to continue, and strengthen and boost appropriate specific responses. Helper T cells have sometimes been called the “conductors” of the immune system because they coordinate activity like the conductor of a symphony. They have also been called the “generals” of the immune system because they call up troops of B cells, cytotoxic T cells, and other helper T cells to go into battle against invading pathogens (Fig. 1).

Figure 1. A specialized macrophage ingests foreign antigens and displays antigen fragments along with MHC (self) molecules on its surface. A helper T cell (TH) with the appropriate receptor binds and responds by producing cytokines that stimulate antigen-specific B cells, as well as specific cytotoxic T cells.
Macrophages alert helper T cells to the presence of pathogens. These phagocytic macrophages engulf bacteria and viruses, and can display foreign antigens — the identifying proteins of the bacteria or viruses — on the surface of their cell membrane. Embedded within the macrophage cell membrane is a molecule produced by the human leukocyte antigen (HLA) complex. (See the Human Evolution unit.) The helper T cells bind simultaneously to the foreign antigen and the HLA molecule. Only TH cells with receptors that match those of the foreign antigen on the activated macrophage are able to bind and respond to the call to action. Once bound, the helper T cell proliferates to form a clone of cells, each capable of recognizing the same antigen. The members of the helper T clone, the generals, generate the chemical signals that call up the troops.
Some signals sent by helper T cells stimulate cytotoxic T cells (TC). Cytotoxic T cells (also known as killer T cells) bind cells that have been altered, such as by viral infection; they avoid healthy cells. Surface antigens on the altered cell perform the binding. These antigens are specific to the offending agent, and match receptors in the membrane of the specific TC cell. In addition, the TC cell simultaneously binds an MHC molecule on the surface of the infected cell. Once bound by both the foreign antigen and the HLA molecule, the cytotoxic T cell secretes a chemical called “perforin,” which destroys the offending cell (Fig. 2).

Figure 2. Binding by both the antigen and an MHC molecule initiates the secretion of lytic enzymes by the cytotoxic T cell (Tc).
Helper T cells also stimulate the production of antibodies. Chemical signals from helper T cells stimulate the production of B cells specific to an infecting pathogen, and then stimulate the B cells to differentiate into plasma cells. The plasma cells are factories for the production of antibodies, which are specific to given pathogens circulating in blood or lymph. Antibodies work by blocking the receptors that allow pathogens to attach to target cells, or by creating clumps of bacteria. Clumping makes the job of phagocytes easier, as they will more readily engulf bacteria in clumps. Bound antibodies sometimes serve as tags, called opsonins, enhancing phagocytosis. Antibody binding can also initiate a cascade of biochemical reactions, activating a set of chemicals known as complement. Activated complement components can form holes in bacterial membranes and enhance inflammation.
Helper T cells are clearly critical to the operation of the immune system. If they are destroyed because of an HIV infection, the whole system is crippled. The immune system is described as having two “arms”: the cellular arm, which depends on T cells to mediate attacks on virally infected or cancerous cells; and the humoral arm, which depends on antibodies to clear antigens circulating in blood and lymph. As an HIV infection progresses, destroying helper T cells, both arms of immunity are impaired.
4. The Structure and Life Cycle of HIV
How does HIV evade the immune system so efficiently? Why are so many variants of the virus found in a single patient? Understanding the structure and life cycle of the virus is key to answering these questions and essential to the design of effective treatments.

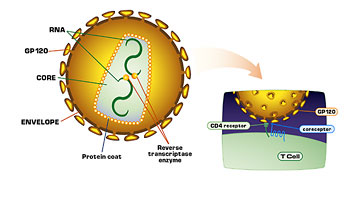
Figure 3. Shows the binding of HIV to a host cell. gp120 on the virus binds CD4 receptors on the host. A second coreceptor molecule on the host is also required for binding.
HIV is an enveloped RNA virus: As HIV buds out of the host cell during replication, it acquires a phospholipid envelope. Protruding from the envelope are peg-like structures that the viral RNA encodes. Each peg consists of three or four gp41 glycoproteins (the stem), capped with three or four gp120 glycoproteins. Inside the envelope the bullet-shaped nucleocapsid of the virus is composed of protein, and surrounds two single strands of RNA. Three enzymes important to the virus’s life cycle — reverse transcriptase, integrase, and protease — are also within the nucleocapsid (Fig. 3).
Although helper T cells seem to be the main target for HIV, other cells can become infected as well. These include monocytes and macrophages, which can hold large numbers of viruses within themselves without being killed. Some T cells harbor similar reservoirs of the virus.
Entry of HIV into the host cell requires the binding of one or more gp120 molecules on the virus to CD4 molecules on the host cell’s surface. Binding to a second receptor is also required. Ed Berger helped identify this coreceptor. As he compared his results with those of other researchers, it became clear that two different coreceptors are involved in the binding. One, CCR5, a chemokine receptor, serves as a coreceptor early in an infection. Another chemokine receptor (CXCR4) later serves as a coreceptor. That two coreceptors are involved is consistent with previous observations. Viruses isolated from individuals early in an infection, during the asymptomatic phase, will typically infect macrophages in the laboratory, but not T cells (the viruses are M-tropic). Virus isolated from patients later in the infection in the symptomatic phase, will infect T cells (the viruses are T-tropic). It seems that a shift takes place in the viral population during the progression of the infection, so that new cellular receptors are used and different cells become infected.

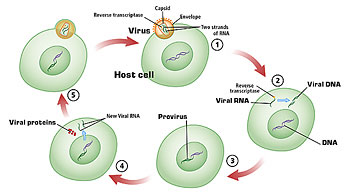
Figure 4. 1) Membranes of the virus and the host cell fuse, and viral RNA and reverse transcriptase enter the host’s cytoplasm. 2) Reverse transcriptase allows viral RNA to be copied to DNA. 3) Viral DNA is incorporated into the host chromosome as provirus. 4) Transcription and translation of viral proteins: viral RNA becomes incorporated into viral particles and is transcribed as well. 5) Viral particles bud out of the host cell, acquiring an envelope in the process.
HIV is a member of the group of viruses known as retroviruses, which share a unique life cycle (Fig. 4). Once HIV binds to a host cell, the viral envelope fuses with the cell membrane, and the virus’s RNA and enzymes enter the cytoplasm. HIV, like other retroviruses, contains an enzyme called reverse transcriptase. This allows the single-stranded RNA of the virus to be copied and double-stranded DNA (dsDNA) to be generated. The enzyme integrase then facilitates the integration of this viral DNA into the cellular chromosome. Provirus (HIV DNA) is replicated along with the chromosome when the cell divides. The integration of provirus into the host DNA provides the latency that enables the virus to evade host responses so effectively.
Production of viral proteins and RNA takes place when the provirus is transcribed. Viral proteins are then assembled using the host cell’s protein-making machinery. The virus’s protease enzyme allows for the processing of newly translated polypeptides into the proteins, which are then ultimately assembled into viral particles. The virus eventually buds out of the cell. A cell infected with a retrovirus does not necessarily lyse the cell when viral replication takes place; rather, many viral particles can bud out of a cell over the course of time.
HIV Transmission
HIV is transmitted principally in three ways: by sexual contact, by blood (through transfusion, blood products, or contaminated needles), or by passage from mother to child. Although homosexual contact remains a major source of HIV within the United States, “heterosexual transmission is the most important means of HIV spread worldwide today.” 2 Treatment of blood products and donor screening has essentially eliminated the risk of HIV from contaminated blood products in developed countries, but its spread continues among intravenous drug users who share needles. In developing countries, contaminated blood and contaminated needles remain important means of infection. Thirteen to thirty-five percent of pregnant women infected with HIV will pass the infection on to their babies; transmission occurs in utero, as well as during birth. Breast milk from infected mothers has been shown to contain high levels of the virus also. HIV is not spread by the fecal-oral route; aerosols; insects; or casual contact, such as sharing household items or hugging. The risk to health care workers is primarily from direct inoculation by needle sticks. Although saliva can contain small quantities of the virus, the virus cannot be spread by kissing.
5. Progression of HIV Infection
Characteristically, an HIV infection can progress for eight to ten years before the clinical syndrome (AIDS) occurs. The long latent period of the virus has contributed to many of the problems relating to diagnosis and control. The basketball player Magic Johnson was still relatively healthy twelve years after he announced he had HIV. On the other hand, not all cases exhibit the long latent period, and abrupt progression to AIDS occurs. Many factors, including genetics, determine the speed at which the disease will progress in a given individual.

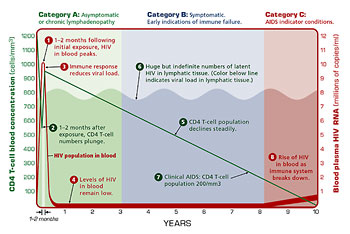
Figure 5. Graph of the typical progression of HIV infection and AIDS.
The Centers for Disease Control and Prevention (CDC) has identified the stages of a typical HIV infection: Categories A, B, and C. In the first stage, Category A, it can be difficult to determine whether an individual is infected without performing a blood test. While at least half of infected individuals will develop a mononucleosis-like illness (headache, muscle ache, sore throat, fever, and swollen lymph nodes) within three weeks of exposure, some Category A individuals are asymptomatic. Moreover, the symptoms themselves can be the result of many different infections. The presence of a rash may help differentiate an HIV infection from other infections, but not all HIV-infected individuals get a rash. Most of these signs and symptoms subside, but swollen lymph glands and malaise can persist for years through Category A HIV.
The number of virus particles circulating in the bloodstream is usually highest soon after exposure. At this point the CD4 cell population plunges (helper T cells are among the immune cells that express the CD4 receptor, which can be used as a marker for counting cell types). As antibodies to HIV appear the numbers of CD4 cells rise; however, CD4 cell levels drop again as the infection progresses. This lowering of CD4 cell levels typically happens slowly, over the course of years. Category C HIV (clinical AIDS) occurs once CD4 numbers have fallen substantially (to 200/mm3 from the normal level of 800-1200 cells/ mm3).
In the Category B stage indications of immune system failure begin. Persistent infections – such as yeast infections, shingles, diarrhea, and certain cancerous conditions of the cervix — are apparent.
Category C is synonymous with AIDS. In this stage the opportunistic infections associated with AIDS appear. According to the CDC, twenty-six known clinical conditions affect people with AIDS; most are infections that do not usually affect healthy individuals. These include yeast infections of the esophagus, bronchi, and lungs; Pneumocystis pneumonia (a fungal infection); toxoplasmosis (caused by a protozoan that is spread by cats); Kaposi’s sarcoma (a rare cancer of the skin caused by a virus); cytomegalovirus (CMV) infections; and tuberculosis. In addition, individuals who have been affected by HIV are more likely to become seriously ill or die than other members of the population during outbreaks of infections such as cryptosporidium (a water-borne parasite) and coccidiomycosis (a dust-borne fungus).
Cytomegalovirus (CMV) causes another opportunistic infection prevalent in AIDS patients. About eighty percent of people in the U.S. have antibodies to this virus, but infections in normal individuals often go undetected or seem like a mild case of mononucleosis. In the immunocompromised, however, CMV can cause life-threatening pneumonia or encephalitis. In AIDS patients CMV that has been latent can reactivate and sometimes cause retinitis, affecting eyesight.
Tuberculosis (caused by Mycobacterium tuberculosis) has been on the rise in the wake of AIDS, such that some call it a co-epidemic. M. tuberculosis causes a respiratory infection (formerly called consumption) that is spread by inhalation. As a result, unlike HIV, behavior modification is less likely to reduce one’s chances of exposure. The bacteria, which have an unusually waxy cell wall, survive well in the environment. M. tuberculosis reproduces inside macrophages found in the lung, and stimulates the production of aggregates of immune cells and connective tissue, called tubercles. Viable organisms can be walled off within such structures for decades, only to become reactivated when a person becomes compromised. Most tuberculosis in AIDS patients results from reactivated infections. AIDS patients suffer not only from respiratory infection but also from disseminated tuberculosis, which can involve the lymphatic system, peritoneum, meninges, urogenital system, or digestive tract. Antibiotic-resistant mycobacteria are also contributing to the rise of tuberculosis, so that second- and third-line drugs must often be used. And because treatments are prolonged, lasting as long as a year, patients sometimes do not complete therapy appropriately. Mycobacteria other than M. tuberculosis, particularly M. avium-intracellulare (MAC), also affect AIDS patients.
Why Do Some Individuals Never Get AIDS?
Despite repeated exposure, some individuals never become infected with HIV. These individuals often have unusual helper T cells with a less-efficient variant of the coreceptor CCR5, which is necessary for viral entry into helper T cells. (See the Human Evolution unit.)
There are also individuals who become infected, but do not progress to AIDS. These long-term survivors, or long-term non-progressors, include individuals who have been AIDS-free as long as eighteen years after infection. A variety of factors may be responsible; for example, infection with less-virulent viruses. Some long-term non-progressors seem to have CD8 cells, which are particularly adept at curtailing HIV infection. (In most AIDS patients CD8 cells become less active.) Several investigators, including Jay Levy (University of California, San Francisco), are evaluating the CD8 cells of long-term survivors to see of they secrete an antiviral protein or proteins that may act against HIV.
Genetic Variation Among HIV
There are five major subtypes of HIV, designated A through E. Different subtypes predominate in different geographical areas. For example, subtype B is more common in North America. In contrast, subtype C predominates in sub-Saharan Africa. Considerable variation within a given subtype also exists. In fact, any given individual infected with HIV will harbor multiple variants of the virus. HIV makes many mistakes as is copies its viral RNA to the DNA that integrates into the host’s chromosome. Because of its sloppy copying of reverse transcriptase, HIV’s mutation rate is high, causing great variability. This large number of variants makes the virus more difficult to treat and hinders vaccine development. In addition, because of its rapid rate of evolution, even within a single individual, HIV can quickly evolve resistance to the drugs the individual is taking to combat the virus.
6. Treatments Based on Understanding the Viral Life Cycle
Treating viruses is always difficult because viruses use the translational machinery of the host cell. Most drugs that target the virus also damage the host. Drugs that can inhibit enzymes specific to the virus are, therefore, less likely to cause side effects in the host.
Most common anti-HIV drugs block key steps in viral reproduction and uptake. Several anti-retroviral drugs work by interfering with reverse transcriptase, the key enzyme of retroviruses. These drugs, the reverse transcriptase inhibitors, act when cells first become infected. Included in this group are the nucleoside analogs, chemicals that are similar to one of the bases (adenine, cytosine, guanine, and thymine) that comprise DNA, but sufficiently different enough to block viral DNA synthesis. There are also non-nucleoside reverse transcriptase inhibitors that can bind to reverse transcriptase and, thus, block the production of viral DNA. Reverse transcriptase inhibitors have been remarkably successful in preventing the spread of HIV from an infected mother to her newborn: if a pregnant woman treated with AZT (a nucleoside analog) delivers her child by caesarian, the chances of the baby being infected can be reduced to one percent.
Protease inhibitors, another major class of drugs, act later in the life cycle of the virus by inhibiting the protease enzyme. These drugs interfere with the cleavage of the viral polypeptide into functional viral enzymes.
The evolution of HIV variants that are resistant to the more commonly used medications has become a major problem. In one study as many as thirty percent of HIV patients harbored resistant viruses. The virus mutates rapidly, and variants that are able to survive in the presence of drug – particularly when circulating levels of the drug are lower – rapidly take over the population. Patient adherence to drug regimens is critical to reducing the emergence of resistant viruses; even the timing of medication can be important. Unfortunately, given the side effects of current treatments, adherence is difficult. Protease inhibitors can cause nausea and diarrhea, and some of the nucleoside reverse transcriptase inhibitors can cause red or white blood cell levels to drop. Painful nerve damage and inflammation of the pancreas can also result.
HAART
Beginning in the mid-1990s, an increasing number of HIV-infected individuals began a drug regime called highly active antiretroviral therapy (HAART), a combination of three or more anti-HIV drugs taken at the same time. The simultaneous intake of multiple drugs, each targeting different aspects of the viral life cycle, circumvents the ability of the virus to mutate and become resistant to the drugs. Combined therapies, often called “cocktails,” can knock virus back to undetectable levels and improve patient health significantly. With the advent of HAART, deaths from HIV began to decline in the U.S. in 1997. Unfortunately, HAART has several long-term side effects including kidney, liver, and pancreatic problems; and changes in fat metabolism, which result in elevated cholesterol and triglyceride levels and an increased risk for strokes and heart attacks. In addition, some viruses have evolved resistance to HAART. Given these side effects, some physicians recommend that HAART be delayed until HIV-positive patients are exhibiting clear signs of AIDS. Still, HAART is often recommended in the first few weeks after exposure to bring the initial viral load down.
The treatments described above are directed at the reduction of free virus: they work only against viruses that are actively produced. Because of the latent nature of the virus they are not cures. In addition, treatments are prolonged and may be necessary for a patient’s entire life. A patient who stops treatment will typically have an increase in viral numbers.
Also under investigation are treatments that take advantage of our understanding of the process of viral infection. “Fusion” or “entry inhibitors” block the proteins involved in viral uptake, such as CCR5. Integrase inhibitors affect the enzyme necessary for the integration of viral DNA into host DNA. Both have shown promise.
Treatments Based on Understanding the Immune System
Development of novel treatments for HIV also depends on an understanding of the choreography of chemical signals that regulate immune function. Because cellular immunity is key to clearing viral infections, increasing the T cell response is critical to clearing HIV. Interleukin 2 (IL-2) is a cytokine produced by TH cells that promotes the growth of other T cells. Recombinant IL-2, which has the same activity as the native protein, has been shown to increase CD4 cell numbers in individuals in the early stages of HIV infection. Viral numbers, though, do not seem to go down with this treatment alone. However, IL-2 administered with HAART resulted in more individuals with undetectable viral loads when compared to treatment with HAART alone. One frustration with HIV treatments is the inability to affect cells that harbor provirus. IL-2 administered intermittently to patients with more advanced HIV could work to stimulate viral production and stimulate HIV specific immune responses. Such strategies are under investigation.
Other treatments under consideration target virally infected cells. Some CD8 cells seem to secrete soluble factors that suppress HIV replication. Understanding how these factors work may help define new treatments.
7. The Challenges of Vaccine Development
Scientists have taken a number of approaches to the development of a vaccine for HIV, but the nature of the virus presents significant challenges. HIV infects only humans and chimpanzees. Evaluating vaccine effectiveness in the chimpanzee model is problematic for several reasons. Chimpanzees are scarce, expensive, and do not show signs of disease when infected. There are also ethical concerns raised because chimpanzees are our closest evolutionary relatives. An alternative is the development of a monkey model using simian immunodeficiency virus (SIV) that has been genetically engineered to express HIV components. The downside to this approach is the difficulty of predicting what will happen when a vaccine that was developed using monkey models is administered to humans.
The route of transmission of HIV also presents a challenge for vaccine producers. Typically, an individual is exposed to the virus at a mucosal surface where a particular type of antibody molecule, IgA, mediates immunity. The ideal vaccine should stimulate production of this type of antibody, not just the type found in the circulation (IgG). But even if a vaccine stimulates the production of the appropriate type of antibody, an increasing number of investigators are convinced that it may not be enough. Circulating antibodies cannot clear a latent virus, and infected cells seem to persist in the body for long periods. So it may be necessary to stimulate cellular as well as humoral immunity. Another challenge to vaccine production is the variety of viral subtypes. Because distinct HIV subtypes are more prevalent in certain locations, some scientists have asked whether HIV vaccines need to be developed specifically for certain geographical regions. Alternately, immune stimulation must be accomplished using an antigen, or antigens, common to all subtypes.
Another major impediment to vaccine development is HIV’s rapid mutation rate and the presence of multiple viral variants within a given individual. Traditional vaccines, such as those for childhood illnesses, consist of live attenuated (weakened) pathogens, dead pathogens, or parts of organisms. Attenuated HIV vaccines are not likely to be pursued because of the risk of infection — whole, killed HIV is a safer alternative. But, given the rapid mutation rate of the virus, many believe that a variant of the virus unaffected by the immune response would evolve quickly.
Vaccines based on pieces of HIV are safer and easier to prepare. Many efforts have been directed to the production of recombinant HIV proteins that can serve as vaccines. For example, vaccines consisting of the gp120 surface protein, which is needed for virus to adhere to cells, could elicit an immune response, inhibiting viral adherence. Unfortunately, gp120 vaccines may not be successful: the site on gp120 that binds CD4 and CCR5 is apparently buried in a molecular pocket, which is not blocked by antibody.
The AIDS epidemic has spurred additional vaccine production strategies that use genetic engineering techniques. Many scientists are examining strategies for generating cellular and humoral immunity; for example, live non-pathogenic bacteria or viruses can be engineered to express HIV antigens. Researchers at Merck Corporation have inserted the gag gene, which encodes a viral core protein, into modified adenovirus. They hope that as cell-mediated immunity is mounted against adenovirus, the response will also target HIV-infected cells. The protein encoded by gag is among those found unchanged in most HIV variants; therefore, researchers hope that the vaccine could circumvent the genetic variability problem.
The gag gene is also the basis of one of several DNA-based vaccines under investigation. Such vaccines contain “naked” DNA (not associated with chromosomes or other structures), which is injected directly into muscle tissue. The expectation is that some of the DNA will be taken up and expressed by human cells. The immune response directed against these cells is hoped to carry over to HIV-infected cells. Some investigators are combining strategies; for example, Harriet Robinson and her colleagues at Emory University are trying DNA priming, followed by a booster of recombinant pox virus.
Clinical Trials
More than two dozen experimental HIV vaccines are being studied worldwide. For a given vaccine to be proven safe and effective it must pass through three stages of human testing. Phase I addresses safety and dosage, and involved the administration of the vaccine to dozens of people. Phase II examines efficacy, the ability of the vaccine to elicit an immune response, and involves hundreds of people. Phase III involved thousands of people who are followed for long periods to establish that the vaccine is indeed protective.
At the outset of the AIDS epidemic some scientists anticipated the availability of a vaccine in two or three years. More than twenty years into the epidemic, a vaccine is still down the road and few believe it will be available soon. The idea of distributing a less-than-perfect vaccine is controversial. Some believe protecting only a certain percentage of the population could limit the spread of the disease. Others believe an imperfect vaccine could provide a false sense of security such that individuals might increase risky behaviors.
8. Social Obstacles to Controlling HIV
Researchers have worked diligently and gained an unprecedented knowledge of the biology of HIV and its interaction with the immune system; yet, the AIDS pandemic will continue for years to come. Obstacles to AIDS prevention and control lie not only in the nature of the HIV virus but the very nature of human societies worldwide. Poverty and discrimination exclude those most in need from information and treatment. The control of HIV lies not only in biology but also in the social realm of basic human rights.
AIDS is having the greatest impact in countries ridden with poverty, where public health infrastructures are already strained by drug-resistant malaria, tuberculosis, yellow fever, Rift Valley fever, and other infectious diseases. (See the Emerging Infectious Diseases unit.) Further, the presence of HIV amplifies epidemics of such pathogens. AIDS is the leading cause of death in Africa. In several African countries, more than twenty percent of the 15-49-year-old population is infected with HIV; in Botswana more than thirty percent of that age group is infected. Poverty excludes millions from treatment. Of the roughly 28 million people infected with HIV in sub-Saharan Africa, only 36,000 received drugs in 2002. In response to such statistics drug companies have reduced the cost of treatment to as little as $300-$400 per person in developing countries (treating one person costs at least $10,0000 or more annually in the U.S.) — but even that is too expensive. In 2001 the United Nations launched the Global Fund to Fight HIV, Tuberculosis, and Malaria. At the time Kofi Annan, U.N. secretary-general, said it would take $7 billion to $10 billion each year to fight HIV/AIDS. As of 2002 the fund, supported mainly by donor nations and philanthropists, had raised only $2 billion.
Poverty is just one obstacle to controlling HIV. Discrimination against particular groups has hindered education, diagnosis, and treatment. The lack of women’s rights in some countries has thwarted educational efforts and contributed to the spread of the disease; so has prevailing customs regarding multiple sex partners. Access to basic education, information about HIV transmission, and the power to say no to unwanted sexual advances are as important as access to drugs. Funding for teacher training, education, and prevention materials have been inadequate.
Governments in many countries have been hesitant to implement strong and coordinated AIDS prevention programs. Needle exchange programs for drug users, for example, have been shown in numerous studies to reduce the risk of HIV. Yet, in countries around the world, such programs remain politically unpopular. Condoms protect against transmission of the virus, but promotions of condom use are discouraged by many religious groups and governments. Children around the world are denied access to sex education, mostly for ideological reasons.
By depleting the workforce, AIDS is destabilizing the economies of countries already grappling with poverty and political instability. As people in their twenties and thirties die, countries lose their workers, their teachers, and the parents of their children. Men who have gone to urban areas to work contract HIV, and then return home to give the disease to their wives. Much of the toll of AIDS in Africa is on the women and children, who are critical to maintaining the continent’s agricultural economy. In many sub-Saharan countries, women are considered their husband’s property and have little access to independent income. As men are lost to AIDS their widows become dependent on others or turn to one of the few survival strategies, prostitution. So viral dissemination is amplified and, at the same time, urban and rural economies decline. By the year 2011 there will be 40 million AIDS orphans in Africa. In some countries the illness and death of women in the childbearing years will result in a greatly reduced number of births in the next decades.
In industrialized societies, those touched by AIDS have had considerable impact in fighting the spread of infection. Patients themselves have become active channels for the distribution of information, and participate in policy-making and lobbying for funding. But HIV remains latent in those whose voices are repressed. In sub-Saharan Africa, for example, stigma still surrounds people infected with HIV, and silence impedes progress in controlling the epidemic.
HIV is difficult to control because it exploits the immune system designed to stop it and other infections. Researchers are continuing to explore strategies to foil the virus, but treatments and vaccines are just two components to thwarting the epidemic. Policies that ensure accessibility to medications and information are critical. Education is key. AIDS forces us to talk about things we would rather leave unsaid.
End Notes
- Coffin, J. M. 1999. Molecular biology of HIV. In The Evolution of HIV, ed. K. A. Crandall, 3–40. Baltimore: Johns Hopkins University Press.
- Amborzia, J., and J. A. Levy. 1998. Epidemiology, natural history and pathogenesis of HIV infection. In Sexually Transmitted Diseases, 3d ed, ed. K.K. Holmes, P.F. Sparling, P.A. Mardh, S.M. Lemon, W.E. Stamm, P. Piot, and J.N. Wasserheit, 251–58. New York: McGraw-Hill.